Митохондриальная днк
Содержание:
- Общие сведения
- Синдром истощения ДНК
- Изменчивость генома
- Структура митохондрий
- Наиболее вероятные нарушения при мутациях митохондрий
- Митохондриальная Ева и африканское происхождение людей
- Формы и число молекул митохондриальной ДНК[править]
- Митохондриальный синдром у детей
- Примечания[править]
- Митохондриальная Ева и африканское происхождение людей
- Структура митохондриальной ДНК
- Синдромы делеции митохондриальной ДНК
Общие сведения
Митохондриальные заболевания обусловлены генетическими, структурными, биохимическими дефектами митохондрий, приводящими к нарушениям тканевого дыхания. Они передаются только по женской линии к детям обоих полов, так как сперматозоиды передают зиготе половину ядерного генома, а яйцеклетка поставляет и вторую половину генома, и митохондрии. Патологические нарушения клеточного энергетического обмена могут проявляться в виде дефектов различных звеньев в цикле Кребса, в дыхательной цепи, процессах бета-окисления и т. д.
Не все ферменты и другие регуляторы, необходимые для эффективного функционирования митохондрий, кодируются митохондриальной ДНК. Большая часть митохондриальных функций контролируется ядерной ДНК.
Можно выделить две группы митохондриальных заболеваний:
Ярко выраженные наследственные синдромы, обусловленные мутациями генов, ответственных за митохондриальные белки (синдром Барта, синдром Кернса-Сейра, синдром Пирсона, синдром MELAS, синдром MERRF и другие).
Вторичные митохондриальные заболевания, включающие нарушение клеточного энергообмена как важное звено формирования патогенеза (болезни соединительной ткани, синдром хронической усталости, гликогеноз, кардиомиопатия, мигрень, печёночная недостаточность, панцитопения, а также гипопаратиреоз, диабет, рахит и другие).
Синдром истощения ДНК
Синдром митохондриального истощения ДНК — весьма редкое заболевание. Ребенок с такой наследственной болезнью — инвалид с детства. Эти синдромы также подразделяются на множество видов.
Многие дети погибают от множественных дефектов развития внутренних органов, не дожив до 3 лет. Получение таких «покалеченных» митохондрий от матери происходит по аутосомно-рецессивному типу наследования. Генетики уверены, что в таких случаях имеют место множественные делеции.
Синдром также называют в научных кругах — синдром истощения митохондриальной ДНК. Заболевание проявляется у новорожденного сразу. У больного младенца наличествуют такие аномалии развития:
- Тяжелая гепатопатия — нарушение работы печени.
- Врожденная миопатия, выраженная в значительной слабости мышц.
- Кардиомиопатия — проблемы в работе сердечной мышцы.
- Атрофия мышц и отсутствие сухожильных рефлексов.
Основная причина таких заболеваний — это дефект межгеномной взаимосвязи (коммуникации).
Существует в генетике и такое понятие как синдром деплеции митохондриальной ДНК. Деплеция — это синоним истощения в генетике. При таком тяжелом синдроме генетический материал митохондрий истощен на 70–98%. Описан впервые не так давно, в 1991 году.
Что происходит с ребенком? В новорожденном периоде уже проявляется лактоацидоз, гипоальбуминемия (резкое снижение альбумина в крови), отеки и выраженная печеночная недостаточность. Наблюдались у некоторых больных и судороги. Симптом, который виден невооруженным взглядом — выраженная мышечная гипотония. Все дети, рожденные с такими признаками, не доживали и до года.
Причиной считается нарушение гена, который ответственен за репликацию ДНК. Его «неверная» работа приводит к тому, что практически все митохондрии мутируют и не выполняют свои функции. Тип наследования делеции митохондриальной ДНК может быть как аутосомно-рецессивным, так и аутосомно-доминантным.
Изменчивость генома
В митохондриальных геномах обнаружено шесть основных типов генома. Эти типы геномов были классифицированы «Колесниковым и Герасимовым (2012)» и различаются различными способами, такими как круговой, по сравнению с линейным геномом, размером генома, наличием интронов или подобных плазмидных структур, а также является ли генетический материал особой молекулой, коллекцией гомогенных или гетерогенных молекул.
Расшифровка генома животных
В клетках животных существует только один тип митохондриального генома. Этот геном содержит одну круговую молекулу между 11-28кбп генетического материала (тип 1).
Расшифровка генома растений
Существует три различных типа генома, содержащихся в растениях и грибах. Первый тип — это круговой геном, который имеет интроны (тип 2) длиной от 19 до 1000 кбп. Второй тип генома представляет собой круговой геном (около 20-1000 кбп), который также имеет плазмидную структуру (1kb) (тип 3). Конечный тип генома, который можно найти в растении и грибах представляет собой линейный геном, состоящий из гомогенных молекул ДНК (тип 5).
Расшифровка генома протистов
Протисты содержат самые разнообразные митохондриальные геномы, которые включают пять разных типов. Тип 2, тип 3 и тип 5, упомянутые в геноме растений и грибов, также существуют в некоторых простейших, а также в двух уникальных типах генома. Первым из них является гетерогенная коллекция круговых молекул ДНК (тип 4), а конечный тип генома, обнаруженный у протистов, представляет собой гетерогенную коллекцию линейных молекул (тип 6). Типы геномов 4 и 6 варьируются от 1 до 200 кб.,
Передача эндосимбиотических генов, процесс генов, кодируемых в митохондриальном геноме, переносится в основном геном клетки, вероятно, это объясняет почему более сложные организмы, например, люди, имеют меньшие митохондриальные геномы, чем более простые организмы, такие как простейшие.
Структура митохондрий
Схема строения митохондрии
Наружная мембрана
Наружная мембрана митохондрии имеет толщину около 7 нм, не образует впячиваний и складок и замкнута сама на себя. На наружную мембрану приходится около 7 % площади поверхности всех мембран клеточных органелл. Основная функция — отграничение митохондрии от цитоплазмы. Наружная мембрана митохондрии состоит из липидов с вкраплениями белков (соотношение 2 : 1). Особую роль играет порин — каналообразующий белок. Он образует в наружной мембране отверстия диаметром 2-3 нм, через которые могут проникать небольшие молекулы и ионы весом до 5 кДа. Крупные молекулы могут проникать сквозь наружную мембрану только посредством активного транспорта с помощью транспортных белков митохондриальных мембран. Для наружной мембраны характерно присутствие ферментов: монооксигеназы, ацил-СоА-синтетазы и фосфолипазы А2. Наружная мембрана митохондрии может взаимодействовать с мембраной эндоплазматического ретикулума; это играет важную роль в транспортировке липидов и ионов кальция.
Межмембранное пространство
Межмембранное пространство представляет собой пространство между наружной и внутренней мембранами митохондрии. Его размер — 10-20 нм. Так как наружная мембрана митохондрии проницаема для небольших молекул и ионов, их концентрация в периплазматическом пространстве мало отличается от таковой в цитоплазме. Крупным же белкам для транспорта из цитоплазмы в периплазматическое пространство необходимо иметь специфические сигнальные пептиды; поэтому белковые компоненты периплазматического пространства и цитоплазмы различны. Одним из белков, содержащихся не только во внутренней мембране, но и в периплазматическом пространстве, является цитохром c.
Внутренняя мембрана
Внутренняя мембрана состоит в основном из белковых комплексов (соотношение белок/липид — 3:1) и образует многочисленные гребневидные складки — кристы, существенно увеличивающие площадь её поверхности и, например, в клетках печени составляет около трети всех клеточных мембран. Характерной чертой состава внутренней мембраны митохондрий является присутствие в ней кардиолипина — особого фосфолипида, содержащего сразу четыре жирные кислоты и делающего мембрану абсолютно непроницаемой для протонов. Ещё одна особенность внутренней мембраны митохондрий — очень высокое содержание белков (до 70 % по весу), представленных транспортными белками, ферментами дыхательной цепи, а также крупными АТФ-синтетазными комплексами. Внутренняя мембрана митохондрии в отличие от внешней не имеет специальных отверстий для транспорта мелких молекул и ионов; на ней, на стороне, обращённой к матриксу, располагаются особые молекулы АТФ-синтазы, состоящие из головки, ножки и основания. При прохождении через них протонов происходит синтез АТФ. В основании частиц, заполняя собой всю толщу мембраны, располагаются компоненты дыхательной цепи. Наружная и внутренняя мембраны в некоторых местах соприкасаются, там находится специальный белок-рецептор, способствующий транспорту митохондриальных белков, закодированных в ядре, в матрикс митохондрии.
Матрикс
Матрикс — ограниченное внутренней мембраной пространство. В матриксе (розовом веществе) митохондрии находятся ферментные системы окисления пирувата, жирных кислот, а также ферменты цикла трикарбоновых кислот (цикла Кребса). Кроме того, здесь же находится митохондриальная ДНК, РНК и собственный белоксинтезирующий аппарат митохондрии.
Наиболее вероятные нарушения при мутациях митохондрий
Всего на сегодняшний день уже выявлено множество форм заболеваний, которые вызывает мутация митохондрий. Например, когда затрагивается мышечный каркас, диагностируют эпилептические приступы на фоне мышечного недоразвития. Причем мышечная структура не просто повреждена, она выглядит под микроскопом как недоразвитые волокна красного цвета. Мышечная атрофия в таком случае называется митохондриальной миопатией. Это наиболее частое нарушение при митохондриальной недостаточности. Если затронута именно сердечная мышца — кардиомиопатия, обнаружены патологические процессы в головном мозге — регистрируется энцефалопатия.
Что такое синдром митохондриальной энцефалопатии? Синдром диагностируют, когда есть нарушения в генах — tRNAs, MTND1, 4–6, MTCYB. При этом нарушается работа всей нервной системы.
Вместе с энцефалопатией наблюдаются и такой симптом, как лактат-ацидоз — или молочнокислая кома. Это осложнение, при котором молочная кислота начинает попадать в кровь.
Опасны и такие состояния у больных с синдромом митохондриальной недостаточности, как частые и злокачественные мигрени, у детей наблюдаются задержки психического и моторного развития, глухота, атаксия (проблемы с равновесием).
Симптомы не так хорошо изучены, поскольку заболевания, связанные с митохондриями не так давно открыты. Но об известных синдромах, клинические проявления которых пытаются лечить, мы расскажем.
Митохондриальная Ева и африканское происхождение людей
Human migrations and mitochondrial haplogroups.PNG
Поскольку популяционные генетики считают родиной митохондриальной Евы Африку, её иногда называют африканской Евой. При древнейшем разделении предковой популяции людей образовались четыре главных гаплогруппы: L0, L1, L2, L3. Из них первая преобладает у бушменов, вторая — у пигмеев. Две последние также имеются у африканских народов, но только от гаплогруппы L3 происходят макрогруппы М и N, носители которых мигрировали из Африки в Евразию.
Существуют альтернативные объяснения митохондриальной генеалогии народов Земли. Например, аналогичное генеалогическое древо могло получиться, если на ранней стадии расселения большинство людей погибло из-за какой-то эпидемии или природной катастрофы, а выжившие представляли собой небольшую группу кровных родственников. Однако попытки построить генеалогию народов на основании исследования других генов не подтверждают гипотезу катастрофы
Кроме того, следует принимать во внимание обмен генами, который происходил в результате браков между дальними родственниками, в результате чего геномы продолжали смешиваться в популяциях уже после исхода из Африки. Наибольшее разнообразие мутаций, которое наблюдается у африканских народов, также можно объяснять по-разному
С одной стороны, это может быть результатом длительного проживания предковой популяции в Африке. С другой стороны, в Африке времён палеолита население могло быть просто многочисленнее, чем в других регионах.
Формы и число молекул митохондриальной ДНК[править]
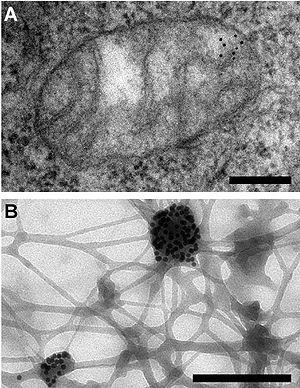
Электронная микроскопия демонстрирует определённую локализацию мтДНК в митохондриях человека. Разрешение 200 нм. (A) Сечение через цитоплазму после окрашивания мтДНК частичками золота. (B) Цитоплазма после экстракции; мтДНК, связанные с частичками золота, остались на месте. Из статьи Iborra et al., 2004.
У большинства изученных организмов митохондрии содержат только кольцевые молекулы ДНК, у некоторых растений одновременно присутствуют и кольцевые, и линейные молекулы, а у ряда протистов (например, инфузорий) имеются только линейные молекулы.
Митохондрии млекопитающих обычно содержат от двух до десяти идентичных копий кольцевых молекул ДНК.
У растений каждая митохондрия содержит несколько молекул ДНК разного размера, которые способны к рекомбинации.
У протистов из отряда кинетопластид (например, у трипаносом) в особом участке митохондрии (кинетопласте) содержится два типа молекул ДНК — идентичные макси-кольца (20-50 штук) длиной около 21 т.п.о. и мини-кольца (20 000 — 55 000 штук, около 300 разновидностей, средняя длина около 1000 п.о.). Все кольца соединены в единую сеть (катенаны), которая разрушается и восстанавливается при каждом цикле репликации. Макси-кольца гомологичны митохондриальной ДНК других организмов. Каждое мини-кольцо содержит четыре сходных консервативных участка и четыре уникальных гипервариабельных участка. В мини-кольцах закодированы короткие молекулы направляющих РНК (guideRNA), которые осуществляют редактирование РНК, транскрибируемых с генов макси-колец.
Митохондриальный синдром у детей
Большинство заболеваний начинается с раннего детского возраста. В основном распространена миопатия, из-за которой дети не могут самостоятельно передвигаться и страдают мышечными болями. Кардиомиопатия — нарушения функции миокарда, также встречается довольно часто.
Митохондриальный синдром у ребенка, если недомогания не слишком серьезны и не угрожают здоровью, на протяжении всей жизни будет причинять беспокойства и мешать нормальному развитию. Таким детям нужны мероприятия по социализации
Им важно развивать скелетные мышцы, но не спортивными методами (так как у многих поражен миокард), а благодаря плаванию с дельфинами. Поэтому создан специальный фонд для таких детей, куда поступают деньги от благотворительности

Однако синдром митохондриального истощения ДНК однозначно приводит к летальному исходу. Врачи предупреждают об этом родителей сразу после установления диагноза. Множественные поражения органов и систем исключают нормальную жизнь для таких детей
Поэтому исключительно важно женщине перед беременностью пройти генетический анализ на мутации в митохондриях
Примечания[править]
- Nass, M.M. & Nass, S. (1963 at the Wenner-Gren Institute for Experimental Biology, Stockholm University, Stockholm, Sweden): Intramitochondrial Fibers with DNA characteristics (PDF). In: J. Cell. Biol. Bd. 19, S. 593—629. PMID 14086138
- Ellen Haslbrunner, Hans Tuppy and Gottfried Schatz (1964 at the Institut for Biochemistry at the Medical Faculty of the University of Vienna in Vienna, Austria): «Deoxyribonucleic Acid Associated with Yeast Mitochondria» (PDF) Biochem. Biophys. Res. Commun. 15, 127—132.
- Iborra FJ, Kimura H, Cook PR (2004). «The functional organization of mitochondrial genomes in human cells». BMC Biol. 2: 9. DOI:10.1186/1741-7007-2-9. PMID 15157274.
- Дымшиц Г. М. Сюрпризы митохондриального генома. Природа, 2002, N 6
- Wiesner RJ, Ruegg JC, Morano I (1992). «Counting target molecules by exponential polymerase chain reaction, copy number of mitochondrial DNA in rat tissues». Biochim Biophys Acta. 183: 553–559. PMID 1550563.
- http://gemi.mpl.ird.fr/PDF/jt01.pdf
- Ченцов Ю. С. Общая цитология. — 3-е изд.. — МГУ, 1995. — 384 с. — ISBN 5-211-03055-9о книге
- Sutovsky, P., et. al (Nov. 25, 1999). «Ubiquitin tag for sperm mitochondria». Nature 402: 371–372. DOI:10.1038/46466. PMID 10586873. Discussed in .
- Vilà C, Savolainen P, Maldonado JE, and Amorin IR (13 June 1997). «Multiple and Ancient Origins of the Domestic Dog». Science 276: 1687–1689. DOI:10.1126/science.276.5319.1687. ISSN 0036-8075. PMID 9180076.
- Hoeh WR, Blakley KH, Brown WM (1991). «Heteroplasmy suggests limited biparental inheritance of Mytilus mitochondrial DNA». Science 251: 1488–1490. DOI:10.1126/science.1672472. PMID 1672472.
-
Penman, Danny. Mitochondria can be inherited from both parents, NewScientist.com, 23 August . Проверено 2008-02-05.
- Kondo R, Matsuura ET, Chigusa SI (1992). «Further observation of paternal transmission of Drosophila mitochondrial DNA by PCR selective amplification method». Genet. Res. 59 (2): 81–4. PMID 1628820.
- Meusel MS, Moritz RF (1993). «Transfer of paternal mitochondrial DNA during fertilization of honeybee (Apis mellifera L.) eggs». Curr. Genet. 24 (6): 539–43. DOI:10.1007/BF00351719. PMID 8299176.
- Fontaine, KM, Cooley, JR, Simon, C (2007). «Evidence for paternal leakage in hybrid periodical cicadas (Hemiptera: Magicicada spp.)». PLoS One. 9: e892. DOI:10.1371/journal.pone.0000892.
- Gyllensten U, Wharton D, Josefsson A, Wilson AC (1991). «Paternal inheritance of mitochondrial DNA in mice». Nature 352 (6332): 255–7. DOI:10.1038/352255a0. PMID 1857422.
- Shitara H, Hayashi JI, Takahama S, Kaneda H, Yonekawa H (1998). «Maternal inheritance of mouse mtDNA in interspecific hybrids: segregation of the leaked paternal mtDNA followed by the prevention of subsequent paternal leakage». Genetics 148 (2): 851–7. PMID 9504930.
- Zhao X, Li N, Guo W, et al (2004). «Further evidence for paternal inheritance of mitochondrial DNA in the sheep (Ovis aries)». Heredity 93 (4): 399–403. DOI:10.1038/sj.hdy.6800516. PMID 15266295.
- Steinborn R, Zakhartchenko V, Jelyazkov J, et al (1998). «Composition of parental mitochondrial DNA in cloned bovine embryos». FEBS Lett. 426 (3): 352–6. DOI:10.1016/S0014-5793(98)00350-0. PMID 9600265.
- Schwartz M, Vissing J (2002). «Paternal inheritance of mitochondrial DNA». N. Engl. J. Med. 347 (8): 576–80. DOI:10.1056/NEJMoa020350. PMID 12192017.
- ↑ Айала Ф. Д. Современная генетика. 1987.
- http://chemistry.umeche.maine.edu/CHY431/MitoDNA.html
- MW Gray, BF Lang, R Cedergren, GB Golding, C Lemieux, D Sankoff, M Turmel, N Brossard, E Delage, TG Littlejohn, I Plante, P Rioux, D Saint-Louis, Y Zhu and G Burger (1998). «Genome structure and gene content in protist mitochondrial DNAs». Nucleic Acids Research 26: 865-878.http://nar.oxfordjournals.org/cgi/content/abstract/26/4/865
- http://kv-journal.su/content/rossiyskie-uchyonye-sozdali-dnk-test-pozvolyayushchiy-nayti-koshatinu-v-shaurme
Митохондриальная Ева и африканское происхождение людей
Human migrations and mitochondrial haplogroups.PNG
Поскольку популяционные генетики считают родиной митохондриальной Евы Африку, её иногда называют африканской Евой. При древнейшем разделении предковой популяции людей образовались четыре главных гаплогруппы: L0, L1, L2, L3. Из них первая преобладает у бушменов, вторая — у пигмеев. Две последние также имеются у африканских народов, но только от гаплогруппы L3 происходят макрогруппы М и N, носители которых мигрировали из Африки в Евразию.
Существуют альтернативные объяснения митохондриальной генеалогии народов Земли. Например, аналогичное генеалогическое древо могло получиться, если на ранней стадии расселения большинство людей погибло из-за какой-то эпидемии или природной катастрофы, а выжившие представляли собой небольшую группу кровных родственников. Однако попытки построить генеалогию народов на основании исследования других генов не подтверждают гипотезу катастрофы
Кроме того, следует принимать во внимание обмен генами, который происходил в результате браков между дальними родственниками, в результате чего геномы продолжали смешиваться в популяциях уже после исхода из Африки. Наибольшее разнообразие мутаций, которое наблюдается у африканских народов, также можно объяснять по-разному
С одной стороны, это может быть результатом длительного проживания предковой популяции в Африке. С другой стороны, в Африке времён палеолита население могло быть просто многочисленнее, чем в других регионах.
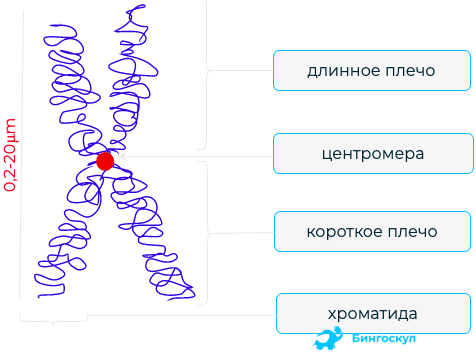
Структура митохондриальной ДНК
В большинстве многоклеточных организмов, мтДНК — или митогеном — организована в виде круглой, циркулярно замкнутой, двухцепочной ДНК. Но во многих одноклеточных (например, тетрахимены или зеленой водоросли Chlamydomonas reinhardtii) и в редких случаях у многоклеточных организмов (например, у некоторых видов книдарий), мтДНК находится как линейно организованная ДНК. Большинство этих линейных мтДНК обладают теломеразо-независимыми теломерами (то есть концами линейной ДНК) с различными режимами репликации, которые сделали их интересными объектами исследования, так как многие из этих одноклеточных организмов с линейной мтДНК являются известными патогенами.
Для митохондриальной ДНК человека (и, вероятно, для метазоанов), 100-10000 отдельных копий мтДНК обычно присутствуют в соматической клетке (яйцеклетки и сперматозоиды являются исключениями). У млекопитающих каждая из двухцепочной молекулы круговой мтДНК состоит из 15000-17000 пар оснований. Две цепи мтДНК различаются по их нуклеотидному содержанию, богатая гуанидом прядь называется тяжелой цепью (или Н-цепью), а богатую цинозином нить называют легкой цепью (или L-нитью). Тяжелая цепь кодирует 28 генов, а легкая — 9 генов, в общей сложности 37 генов. Из 37 генов 13 предназначены для белков (полипептидов), 22 — для передачи РНК (тРНК) и два — для малых и больших субъединиц рибосомальной РНК (рРНК). Митогеном человека содержит перекрывающиеся гены (ATP8 и ATP6, а также ND4L и ND4: см. Карту генома человека митохондрий), которая редко встречается в геномах животных. 37-генная картина также встречается среди большинства метазоанов, хотя, в некоторых случаях, один или несколько из этих генов отсутствуют, а диапазон размеров мтДНК больше. Еще большее изменение содержания и размера генов мтДНК существует среди грибов и растений, хотя, как представляется, существует основное подмножество генов, которое присутствует во всех эукариотах (за исключением немногих, у которых вообще нет митохондрий). Некоторые виды растений имеют огромные мтДНК (столько, сколько 2500000 пар оснований на молекулу мтДНК), но, как ни удивительно, даже эти огромные мтДНК содержат одинаковое число и виды генов, как родственные растения с гораздо меньшими мтДНК.
Геном митохондрии огурца (Cucumis Sativus) состоит из трех кольцевых хромосом (длина 1556, 84 и 45 т.п.н.), которые полностью или в значительной степени автономны в отношении их репликации.
Наименьший митохондриальный геном, упорядоченный до настоящего времени, представляет собой мтДНК 5967bp паразита Plasmodium falciparum; некоторые виды растений имеют огромные митохондриальные геномы с мтДНК Silene conica, содержащей 11,3 Мб.
Синдромы делеции митохондриальной ДНК
Начнем описание с такого заболевания, как синдром Кернса-Сейра, начинающийся с 4 лет. Синдром проявляется следующим образом:
- офтальмоплегия прогрессирующая;
- атаксия;
- атриовентрикулярная блокада сердца (замедление передачи импульса от одной сердечной камеры к другой);
- ретинит пигментный;
- те же красные рваные мышечные ткани.
Следующий синдром, имеющий те же «корни» — синдром Пирсона, который проявляется иначе:
- анемия гипопластическая, самый первый и опасный симптом;
- нарушение функций поджелудочной;
- позже возможны нарушения зрения;
- нарушения в костном мозге;
- появление деменции.
Синдром Пирсона обусловлен, как и синдром Кернса-Сейра, делециями митохондриальной ДНК. Делеции — это такие изменения в хромосомном наборе гена, при которых часть генного материала полностью утеряна.
Те аллели, которые мутировали, или потеряли части хромосом, не должны проявляться как доминантные. Но в митохондриальной ДНК все процессы хаотичны, мутирование происходит слишком быстро. Некоторые ученые даже считают, что митохондрии — это не органеллы, а бактерии, которые когда-то попали в человеческий организм и полностью прижились, создали симбиотическую связь с клеткой и начали служить ей. На такую теорию наталкивает тот факт, что у митохондрии свои, отдельные кольцевые ДНК.