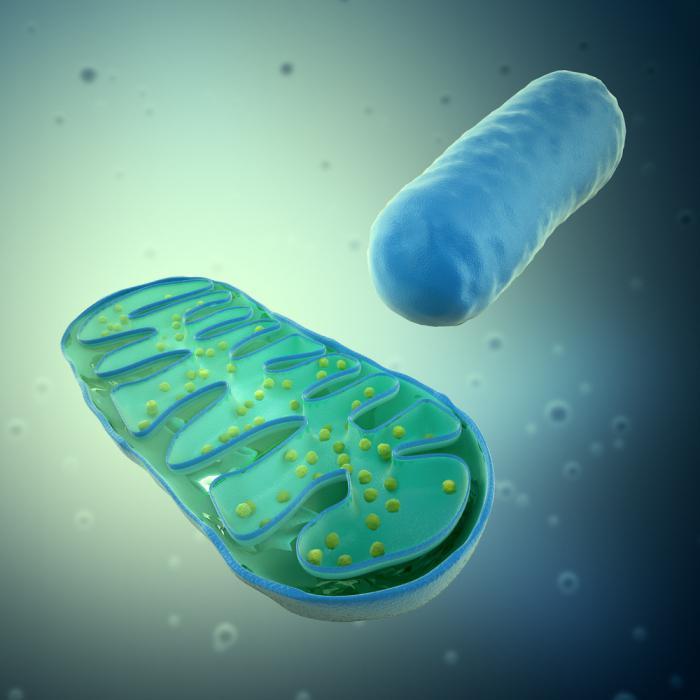
Гаплогруппа t (мтднк)
Содержание:
- Белковая наследственность
- Распространение
- Y‑хромосомный Адам
- Основные понятия и термины генетики
- Распространение
- Примечания
- Геном митохондрий
- Медицина
- Распространение
- Митохондриальная наследственность
- Особенности митохондриальной ДНК[править]
- Палеогенетика
- Наследование митохондриальных болезней
- Палеогенетика
- Митохондрии в клетке
Белковая наследственность
Основная статья: Прионы
Прионы — белковые инфекционные агенты, у человека и других животных вызывают различные нейродегенеративные заболевания. Открытие белковых инфекционных агентов в конце XX века лишь на первый взгляд пошатнуло центральную догму молекулярной биологии. В действительности же прионы не способны к самостоятельной репликации. Прионный белок способен существовать в по меньшей мере двух конформациях: инфекционной и нормальной. Их первичная структура одинакова. Попадая в организм, инфекционный белок укладывает вновь синтезированные гомологичные белки в пространстве по своему образу и подобию. В этом и проявляется их инфекционное начало.
У млекопитающих прионы не передаются по наследству, но у грибов — дрожжей-сахаромицетов и — существует явление прионной (белковой) наследственности. Таким образом, их прионный механизм наследования является наиболее ярким примером собственно цитоплазматической наследственности.
Распространение
Средняя частота гаплогруппы J в целом наиболее высока на Ближнем Востоке (12 %), затем в Европе (11 %), на Кавказе (8 %) и в Северной Африке (6 %). Из двух основных субкладов, J1 составляет 4/5 от всей гаплогруппы и распространена довольно широко, тогда как J2 локализована в Средиземноморье (Греция, Италия/Сардиния и Испания). Некоторые гаплотипы J2 проявляются с заметной частотой в Скандинавии и на Британских островах. В Пакистане, где западноевразийские родословные наблюдаются с частотой до 50 % в ряде этнолингвистических групп, J1 встречается со средней частотой 5 %, тогда как J2 весьма редка. При этом интересно отметить, что J2 встречается с частотой 9 % среди калашей, изолированной этнической группы на Гиндукуше.
В Европе гаплогруппа J встречается в следующих местах (перечислены только те, где её частота превышает 2 %):
- J* = Ирландия — 12 %, Англия и Уэльс — 11 %, Шотландия — 9 %, Оркнейские острова — 8 %, Германия — 7 %, Россия (европейская часть) — 7 %, Исландия — 7 %, Австрия и Швейцария — 5 %, Финляндия и Эстония — 5 %, Испания и Португалия — 4 %, Франция и Италия — 3 %
- J1a = Австрия и Швейцария — 3 %
- J1b1 = Шотландия — 4 %
- J2 = Франция и Италия — 2 %
- J2a = равномерно распределена по Европе. Отсутствует у народов Кавказа. Где-либо в иных местах не представлена.
- J2b1 = практически отсутствует в Европе, однако обнаружена в различных формах на Ближнем Востоке.
Y‑хромосомный Адам
Основная статья: Y-хромосомный Адам
Схожее понятие молекулярной антропологии означает наиболее близкого общего предка всех ныне живущих людей по мужской линии. Так как Y‑хромосома передаётся только от отца к сыну, то все современные Y‑хромосомы происходят от данного мужчины, которого называют Y‑хромосомным Адамом. Подобно тому как митохондриальная Ева не была единственной женщиной в то время, не был единственным и Y‑хромосомный Адам: мужские хромосомы его современников в силу естественного процесса пресечения прямых линий просто не сохранились.
Из-за того, что Y‑хромосома значительно длиннее, чем ДНК митохондрий, примерно на 60 миллионов пар оснований, и имеет более низкую частоту мутаций, идентификация её полиморфизма замедляется и, как следствие, снижается точность оценки частоты мутаций.
В большинстве старых работ возраст Адама оценивался примерно в 100 000 лет и меньше, что создавало забавное несоответствие с оценкой времени жизни Евы в 140 000—200 000 лет назад: таким образом, Ева оказывалась старше Адама не менее чем на 50 000 лет. Вообще говоря, возрасты общих предков по различным участкам ДНК (мтДНК и Y‑хромосомы) не обязаны совпадать, так как процесс исчезновения аллелей в популяции является стохастическим, нет никаких общих закономерностей, требующих «синхронности» эволюции разных локусов. Некоторые учёные даже выдвигали возможные причины такой разницы — из-за практики многожёнства женщины имели больше шансов передать дочерям свои митохондриальные ДНК, чем мужчины сыновьям — Y‑хромосомы: когда у мужчины имеется несколько жён, он фактически устраняет других мужчин от воспроизведения и передачи хромосом в следующие поколения. С другой стороны, многожёнство не мешает женщинам передавать ДНК митохондрий своим детям. Эта разница может привести к уменьшению прямых мужских линий по отношению к женским.
Однако в последний год наметилась стойкая тенденция к «удревнению» Адама в научной литературе: оценки его возраста как по микросателлитным, так и по SNP-данным достигли нижней оценки возраста митохондриальной Евы. Так, Фульвио Кручиани произведены ревизия и диссекция Y‑хромосомной гаплогруппы A, приведшая к смене топологии прикорневой части глобального древа Y‑хромосомы человека и была получена МП-оценка возраста Адама в 142 000 лет. Впрочем недавние исследования обнаружили новую гаплогруппу A00, что значительно отдалило время Y‑хромосомного Адама — от 237 000 до 581 000 лет назад (с вероятностью 95 %).
Тем не менее, в августе 2013 года появились новые данные о том, что Адам жил 120—156 тысяч лет назад (с максимальной вероятностью — 138 тысяч лет назад).
Основные понятия и термины генетики

Основные понятия и термины генетики
Генетика
Генетика (от греч. генетикос – относящийся к происхождению) – это раздел биологии, наука, которая изучает закономерности наследственности и изменчивости организмов. Это название предложил английский ученый У. Бэтсон в 1906 году.
Предметом исследований генетики являются явления наследственности и изменчивости.
Наследственность
Наследственность – это свойство организмов передавать в процессе размножения потомкам способность к определенному типу индивидуального развития и повторять в ряде поколений похожие признаки. Наследственность обеспечивает внутривидовое сходство организмов в ряде поколений.
Изменчивость
Изменчивость – это способность живых организмов приобретать новые признаки во время индивидуального развития. Изменчивость является противоположностью наследственности. Именно она обеспечивает индивидуальное разнообразие особей одного вида.
Изменчивость и наследственность тесно связаны. Новые признаки проявляются лишь благодаря изменчивости и имеют значение в эволюционном процессе видообразования, если наследуются.
Ген
Элементарной единицей наследственности является ген. Ген – это участок молекулы ДНК, который несет информацию о последовательности аминокислот в белке, то есть информацию о наследственном признаке организма.
Под наследственным (генетическим) признаком понимают разные свойства организма, которые передаются потомкам. Генетические признаки делят на качественные и количественные. Качественные: окраска семени, цветка, форма цветка, расположение цветков и т. п. Количественные могут быть измерены. Например: рост, масса, удойность и др.
Альтернативные признаки
Альтернативные признаки – это признаки, проявление которых противоположные, взаимоисключающие, отличные, альтернативные. Например: форма семени – гладкая и морщинистая, окраска цветков – красная и белая и т. д.
Гены в хромосомах расположены в определенном порядке и на определенном месте – локусе. Аллельные гены (аллели) – это гены, расположенные в одинаковых локусах гомологичных хромосом, которые определяют состояние определенного признака. Если состояния одинаковые – диплоидная (полиплоидная) клетка (особь) гомозиготная (от греч. гомос – равный, одинаковый, зиготос – соединенное вместе). Если состояния разные – гетерозиготная (от греч. гетерос – разный).
Фенотип
Совокупность всех свойств и признаков организма называется фенотипом (от греч. файнос – представляю, типос – отражение).
Генотип
Совокупность всех генов, определяющих эти признаки и свойства, называется генотипом (от греч. генос – род). Фенотип формируется вследствие взаимодействия генотипа с внешней средой.
Распространение
Гаплогруппа H — наиболее распространённая митохондриальная гаплогруппа в Европе — к ней относится более половины современного женского населения Северо-Западной Европы. Данная гаплогруппа также часто встречается в Северной Африке и на Ближнем Востоке. Частота распространения данной гаплогруппы в Европе уменьшается к юго-востоку, составляя всего 20 % на Ближнем Востоке и Кавказе, и менее 10 % в Персидском Заливе, Северной Индии и Центральной Азии.
Среди указанных кладов H1 и H3 подверглись наиболее детальному исследованию; их связывают с мадленской экспансией из Юго-Западной Европы около 13 тысяч лет назад:
Подгаплогруппа H1 составляет значительную долю западноевропейских митохондриальных ДНК, причём пик распространения приходится на басков (27,8 %). Также распространена среди других жителей Иберийского полуострова, Северной Африки и Сардинии. Составляет свыше 10 % во многих других регионах Европы (Франция, Британские острова, Альпы, многие регионы Восточной Европы) и не менее 5 % в прочих местах Европы. Субклад H1b имеет наибольшее распространение в Восточной Европы и на северо-западе Сибири.
Подгаплогруппа H3 составляет значительно меньшую долю «общеевропейского генома», чем H1, однако имеет примерно такое же распространение с максимумом среди басков (13,9 %), галисийцев (8,3 %) и сардинцев (8,5 %). Плотность её падает в направлении северо-востока Европы. Ряд исследований показал, что гаплогруппа H3 связана с весьма высокой сопротивляемостью риску заражения СПИДом.
Оставшиеся субклады встречаются намного реже:
Подгаплогруппа H5, вероятно, возникла в Западной Азии, где чаще всего встречается в исходном виде. Её субклад H5a больше всего распространён на Центральноевропейских равнинах.
Подгаплогруппы H2, H6 и H8 довольно часто встречаются в Восточной Европе и на Кавказе. Вероятно, это наиболее распространённые субклады гаплогруппы H среди жителей Средней Азии, изредка встречаются и в Западной Азии.
Подгаплогруппы H4, H7 и H13 присутствуют как в Европе, так и в Западной Азии, а последняя — также на Кавказе. Все три указанных субклада довольно редки.
Примечания
- A. C. Wilson, R. L. Cann, S. M. Carr, M. George Jr., U. B. Gyllensten, K. Helm- Bychowski, R. G. Higuchi, S. R. Palumbi, E. M. Prager, R. D. Sage, and M. Stoneking (1985) «Mitochondrial DNA and two perspectives on evolutionary genetics». Biological Journal of the Linnean Society 26:375-400
- Bryan Sykes The Seven Daughters of Eve: The Science That Reveals Our Genetic Ancestry, W.W. Norton, 2001, hardcover, 306 pages, ISBN 0-393-02018-5
- ↑ Richard Dawkins. The Ancestor’s Tale, A Pilgrimage to the Dawn of Life (англ.). — Boston: Houghton Mifflin Company (англ.)русск., 2004. — ISBN 0-618-00583-8.
- ↑ Soares P, Ermini L, Thomson N, Mormina M, Rito T, Rohl A, Salas A, Oppenheimer S, Macaulay V, Richards MB.Correcting for purifying selection: an improved human mitochondrial molecular clock., Am J Hum Genet 84(6):740-759. 2009
- Поскольку одновременная мутация у нескольких индивидов практически невозможна.
- See the chapter All Africa and her progenies in Richard Dawkins. River Out of Eden (неопр.). — New York: Basic Books, 1995. — ISBN 0-465-06990-8.
- Eva-Liis Loogvali, Toomas Kivisild, Tonu Margus, Richard Villems Explaining the Imperfection of the Molecular Clock of Hominid Mitochondria., PLoS ONE 4(12): e8260, 2009
- Stone et al. // Genes, Culture and Human Evolution (неопр.). — 2007. — ISBN 1-4051-3166-7.
- (primary source)
Геном митохондрий
У млекопитающих каждая молекула мтДНК содержит 15000-17000 пар оснований (у человека 16565 пар нуклеотидов — исследование закончено в 1981 году, по другому источнику 16569 пар) и содержит 37 генов — 13 кодируют белки, 22 — гены тРНК, 2 — рРНК (по одному гену для 12S и 16S рРНК). Другие многоклеточные животные имеют схожий набор митохондриальных генов, хотя некоторые гены могут иногда отсутствовать. Генный состав мтДНК разных видов растений, грибов и особенно протистов различается более значительно. Так, у жгутиконосца-якобиды Reclinomonas americana найден наиболее полный из известных митохондриальных геномов: он содержит 97 генов, в том числе 62 гена, кодирующих белки (27 рибосомальных белков, 23 белка, участвующих в работе электрон-транспортной цепи и в окислительном фосфорилировании, а также субъединицы РНК-полимеразы).
Один из наиболее маленьких митохондриальных геномов имеет малярийный плазмодий (около 6.000 п.о., содержит два гена рРНК и три гена, кодирующих белки).
Недавно открытые рудиментарные митохондрии (митосомы) некоторых протистов (дизентерийной амёбы, микроспоридий и лямблий) не содержат ДНК.
Митохондриальные геномы различных видов грибов содержат от 19 431 (делящиеся дрожжи Schizosaccharomyces pombe) до 100 314 (сордариомицет Podospora anserina) пар нуклеотидов.
Некоторые растения имеют огромные молекулы митохондриальной ДНК (до 25 миллионов пар оснований), при этом содержащие примерно те же гены и в том же количестве, что и меньшие мтДНК. Длина митохондриальной ДНК может широко варьировать даже у растений одного семейства. В митохондриальной ДНК растений имеются некодирующие повторяющиеся последовательности.
Геном человека содержит только по одному промотору на каждую комплементарную цепь ДНК.
Геном митохондрий человека кодирует следующие белки и РНК:
| Белки или РНК | Гены |
| NADH-дегидрогеназа(комплекс I) | MT-ND1, MT-ND2, MT-ND3, MT-ND4, MT-ND4L, MT-ND5, MT-ND6 |
| Кофермент Q — цитохром c редуктаза/Цитохром b(комплекс III) | MT-CYB |
| цитохром c оксидаза(комплекс IV) | MT-CO1, MT-CO2, MT-CO3 |
| АТФ-синтаза | MT-ATP6, MT-ATP8 |
| рРНК | MT-RNR1 (12S), MT-RNR2 (16S) |
| тРНК | MT-TA, MT-TC, MT-TD, MT-TE, MT-TF, MT-TG, MT-TH, MT-TI, MT-TK, MT-TL1, MT-TL2, MT-TM, MT-TN, MT-TP, MT-TQ, MT-TR, MT-TS1, MT-TS2, MT-TT, MT-TV, MT-TW, MT-TY, MT1X |
Медицина
Согласно одному из исследований, результаты которого подверглись критике специалистов, митохондриальная гаплогруппа T связана со сниженной подвижностью сперматозоидов у мужчин.
Согласно публикации кафедры биохимии и молекулярно-клеточной биологии Университета Сарагосы, гаплогруппа T представляет собой слабую генетическую предрасположенность к астенозооспермии.
Согласно некоторым исследованиям, наличие гаплогруппы T связано с повышенным риском коронарно-артериального заболевания. Согласно другому исследованию, носители T менее склонны к диабету.
Несколько пилотных медицинских исследований показали, что наличие гаплогруппы T связано с пониженным риском болезней Паркинсона и Альцгеймера.
Распространение
Встречается в Северо-Восточной Азии (в том числе в Сибири). Также является одной из 5 мтДНК-гаплогрупп, обнаруженных у коренных народов Америки, наряду с такими, как A, B, C и X2a. D1 это базальная ветвь D4, которая широко распространена и разнообразна в Северной и Южной Америке. Субклады D4b1, D4e1 и D4h встречаются как в Азии, так и в Северной и Южной Америке. D2, встречается с высокой частотой в некоторых арктических и субарктических популяциях (особенно у алеутов) и является подклассом D4e1, параллельным D4e1a и D4e1c, поэтому его правильно следует называть D4e1b. D3 был найден главным образом в некоторых сибирских популяциях и у эскимосов Канады и Гренландии и является ветвью D4b1c.
Также гаплогруппа D довольно часто встречается в Центральной Азии, где она является второй по частоте кладой мтДНК (после H). С низкой частотой гаплогруппа D встречается на северо-востоке Европы и юго-Западной Азии.
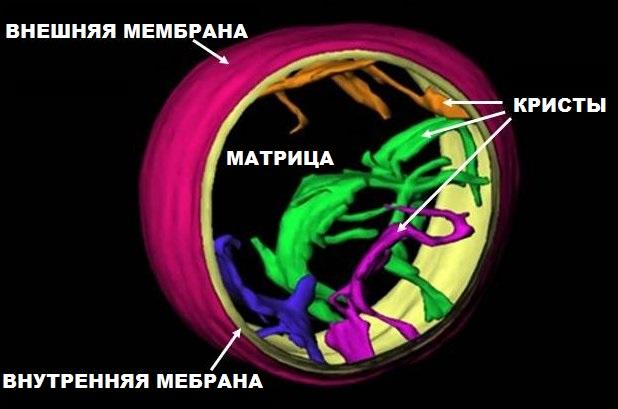
Митохондриальная наследственность
Основные статьи: и Митохондриальные заболевания
Схема митохондриального генома человека
Митохондрии, как и хлоропласты, содержат собственный геном, представленный кольцевой молекулой ДНК. У большинства многоклеточных организмов митохондриальная ДНК наследуется по материнской линии. Это связано, во-первых, с тем, что яйцеклетка содержит во много раз больше митохондрий, чем сперматозоид, и, во-вторых, после оплодотворения митохондрии сперматозоида деградируют. Тем не менее, для некоторых животных описано наследование митохондрий по мужскому типу, например, у мидий, некоторых насекомых; отдельные случаи известны и для млекопитающих. Митохондриальный геном кодирует ряд белков, задействованных в цикле Кребса, β-окислении жирных кислот, и, особенно, окислительном фосфорилировании. Мутации, затрагивающие митохондриальный геном, нередко приводят к развитию различных заболеваний, поскольку они нарушают энергообмен клетки и могут даже привести к её гибели. Несмотря на прогресс в области изучения причин митохондриальных заболеваний, они остаются неизлечимыми и по сей день.
Цитоплазматическая мужская стерильность
Основная статья: Цитоплазматическая мужская стерильность
Цитоплазматическая мужская стерильность — это наследование признаков, ограничивающих или сводящих на нет фертильность мужских растений (например, из-за образования дефектной пыльцы или даже полное её отсутствие, морфологические особенности цветка и т. п.), по материнскому типу через цитоплазму. Следует отметить, что вообще мужская стерильность у растений может определяться и рецессивным аллелем соответствующего ядерного гена. Явление цитоплазматической мужской стерильности описано у более 150 видов растений из 20 различных семейств, в частности, у таких экономически важных видов растений, как кукуруза, пшеница, рожь, сорго, сахарная свёкла, подсолнечник, бобы, морковь, лук.
Цитоплазматическая мужская стерильность обусловлена мутациями мтДНК. Во многих случаях цитоплазматической мужской стерильности наблюдается появление новых химерных генов, образующихся в результате слияния митохондриального гена с какой-либо привнесённой последовательностью из ядерного или хлоропластного генома.
У кукурузы существует особый ядерный ген — восстановитель фертильности (Rf/rf). Находясь в доминантном состоянии, он обеспечивает развитие нормального фертильного растения даже при наличии в цитоплазме фактора стерильности, а рецессивная аллель влияет на репродуктивную функцию при нормальной цитоплазме. Поэтому стерильными будут только растения, гомозиготные по рецессивному аллелю rf и имеющие в цитоплазме фактор стерильности.
У кукурузы (Zea mays) плазмогены (то есть цитоплазматические факторы) мужской стерильности производят плейотропное действие: уменьшается число листьев, снижается устойчивость к некоторым болезням.
Явление восстановления фертильности пыльцы используется на практике для появления гетерозисных двойных межлинейных гибридов кукурузы. Так как кукуруза самосовместима, то, чтобы исключить самоопыление, у некоторых растений приходилось обламывать мужские метёлки, то есть чтобы сделать их исключительно женскими особями. Так что гибриды CytSrf/rf (CytS — стерильная цитоплазма, CytN — нормальная цитоплазма) являются решением этой проблемы, поскольку имеют цитоплазматическую мужскую стерильность и неспособны к самооплодотворению.
Особенности митохондриальной ДНК[править]
В отличие от ядерной ДНК, Мт-ДНК локализуется исключительно в митохондриях. Кодирующие последовательности (кодоны) митохондриального генома имеют некоторые отличия от кодирующих последовательностей универсальной ядерной ДНК.
Так, кодон AUA кодирует в митохондриальном геноме метионин (вместо изолейцина в ядерной ДНК), кодоны AGA и AGG — терминаторные кодоны (в ядерной ДНК кодируют аргинин), кодон UGA в митохондриальном геноме кодирует триптофан.
Если говорить точнее, то речь идёт не о митохондриальной ДНК, а о мРНК, которая списывается (транскрибируется) с этой ДНК перед началом синтеза белка. Буква U в обозначении кодона обозначает уридин, который при транскрипции гена в РНК заменяет тимин.
Количество генов тРНК (22 гена) меньше, чем в ядерном геноме с его 32 генами тРНК.
В человеческом митохондриальном геноме информация настолько сконцентрирована, что в последовательностях кодирующих мРНК, как правило, частично удалены нуклеотиды, соответствующие 3′-концевым терминаторным кодонам.
Палеогенетика
Предполагается, что гаплогруппа T возникла около 33 тыс. лет назад на Ближнем Востоке (в Месопотамии или области Плодородного Полумесяца), после чего примерно 15 тыс. лет назад распространилась в Европе и на восток вплоть до современных Пакистана и Индии.
Гаплогруппа T присутствует в Европе, как предполагается, менее 12 тысяч лет, что делает её самой молодой из европейских митохондриальных гаплогрупп. На сайте Генографического проекта отмечается, что ранние носители гаплогруппы T были, по-видимому, первыми носителями сельскохозяйственной культуры и составляли группу, которая принесла сельское хозяйство в Европу, что стало причиной «неолитической революции».
- Ветвь T2 имеет довольно высокую частоту (20-33 %) среди митохондриальных гаплогрупп неолитических культур центральной Европы (линейно-ленточной керамики и Сакалхат — последняя представляла собой гибрид элементов ЛЛК и культуры Винча). Примерно с аналогичной частотой Т2 (субклад T2b) встречается у носителей трипольской культуры из пещеры Вертеба.
- Субклады T1a и T2b найдены у представителей старчевской культуры, живших ок. 5700 лет назад. Субклада T2b была обнаружена у представителя культуры кардиальной керамики. Субклады T2 и T1 обнаружены у представителей ямной культуры.
- T1a2, T2, T2g1a определены у энеолитических (4500—3900/3800 гг. до н. э.) образцов из израильской пещеры Пкиин (Peqi’in Cave).
- Субклада T2a1a определена у представителя катакомбной культурно-исторической общности. Субклада T2c1a2 была обнаружена у представительницы афанасьевской культуры эпохи бронзы.
- Субклады T1a, T1a2, T1a5, T1a7, T1a8a, T2, T2c1, T2c1c и T2e определены у мумий из Абусира.
- Гаплогруппа T определена у образца из БМАК (Бактрийско-Маргианский археологический комплекс).
- T1 обнаружена у представителя скифской пазырыкской культуры эпохи железного века.
- T2c1a обнаружена у древнеегипетской мумии OM S1 из художественного музея Тартуского университета, датируемой второй половиной 1 тыс. до нашей эры.
- T2 обнаружена у одного из викингов с языческого захоронения Галгедил (Galgedil) на датском острове Фюн (700—1100 гг. н. э.).
- T1, T1a1j, T2 и T2f1 определены у обитателей Сигтуны, живших примерно в X—XII веках.
- T2b21b определена у образца Sk6990 (XVI век) с Серебряного рудника в Швеции.
- T2b была определена у женщины-воина из камерного захоронения Bj.581 на острове Бьёркё, похороненной в X веке в могиле вместе с боевым топором, ножом, копьём, бронебойными стрелами, двумя щитами, жеребцом и кобылой.
- T1 обнаружена у римлянина (ERS3189333, QED-2), с горы Корнет-эд-Дейр (Qornet ed-Deir) на севере Ливана, жившего примерно в 244—400 годах. T2 обнаружена у крестоносца из Сидона (ERS3189343, SI-53), жившего примерно в 1025—1154 годах.
- T1a и T2c1d1 определены у позднетрипольских образцов Pocrovca 3 и Pocrovca 2 (3500—3100 лет до н. э.) из Молдавии, T4 и T2b — у трипольцев из пещеры Вертеба (Тернопольская область Украины).
Наследование митохондриальных болезней
Митохондрии наследуются иначе, чем ядерные гены. Ядерные гены в каждой соматической клетке обычно представлены двумя аллелями (за исключением большинства сцепленных с полом генов у гетерогаметного пола). Один аллель унаследован от отца, другой от матери. Однако митохондрии содержат собственную ДНК, причем в каждой митохондрии человека обычно содержится от 5 до 10 копий кольцевой молекулы ДНК (см. Гетероплазмия), и все митохондрии наследуются от матери. Когда митохондрия делится, копии ДНК случайным образом распределяются между её потомками. Если только одна из исходных молекул ДНК содержит мутацию, в результате случайного распределения такие мутантные молекулы могут накопиться в некоторых митохондриях. Митохондриальная болезнь начинает проявляться в тот момент, когда заметное число митохондрий во многих клетках данной ткани приобретают мутантные копии ДНК (пороговая экспрессия).
Мутации в митохондриальной ДНК происходят, по разным причинам, намного чаще, чем в ядерной. Это означает, что митохондриальные болезни достаточно часто проявляются из-за спонтанных вновь возникающих мутаций. Иногда темп мутирования увеличивается из-за мутаций в ядерных генах, кодирующих ферменты, которые контролируют репликацию ДНК митохондрий.
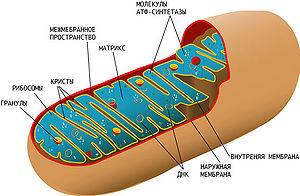
Схема строения митохондрии. Сложная структура митохондрии и наличие собственной кольцевой хромосомы, кодирующей некоторые компоненты митохондрии, усложняет выяснение причин митохондриальных заболеваний
Дефекты и симптомы
Эффекты митохондриальных заболеваний очень разнообразны. Из-за различного распределения дефектных митохондрий в разных органах мутация у одного человека может привести к заболеванию печени, а у другого — к заболеванию мозга. Величина проявления дефекта может быть большой или малой, и она может существенно изменяться, медленно нарастая во времени. Некоторые небольшие дефекты приводят лишь к неспособности пациента выдерживать физическую нагрузку, соответствующую его возрасту, и не сопровождаются серьёзными болезненными проявлениями. Другие дефекты могут быть более опасны, приводя к серьёзной патологии.
В общем случае митохондриальные заболевания проявляются сильнее при локализации дефектных митохондрий в мышцах, мозге, нервной ткани, поскольку эти органы требуют больше всего энергии для выполнения соответствующих функций.
Несмотря на то, что протекание митохондриальных заболеваний сильно отличаются у разных пациентов, на основании общих симптомов и конкретных мутаций, вызывающих болезнь, выделено несколько основных классов этих заболеваний.
Палеогенетика
Древняя митохондриальная ДНК останков, известных как «Пальиччи 23» (en:Paglicci 23) возрастом 28 тысяч лет из пещеры Пальиччи (Апулия, Италия) соответствует эталонной кембриджской последовательности HVR1, указывающей на то, что индивидуум имел либо митохондриальную гаплогруппу R, либо митохондриальную гаплогруппу H. Свидетельством достоверности данного открытия является то, что гаплотип данных останков отличался от гаплотипа всех лиц, работавших с данными останками со времени их обнаружения.
Субклада H13c была обнаружена у мезолитического охотника из карстового грота Котиас Клде в известняках плато Мандаэти в Западной Грузии, жившего 9,529-9,895 тыс. лет назад.
Гаплогруппа H была обнаружена у представителя старчевской культуры, жившего ок. 7600 лет назад и у представителей днепро-донецкой культуры, живших ок. 7500 лет назад.
Субклады H3 и H4a1 были обнаружены у представителей культуры кардиальной керамики, живших 7400 лет назад.
Носителем митохондриальной гаплогруппы H1 был обитатель раннего неолита (7200 — 7000 л. н.) из Карсдорфа (Саксония-Анхальт, Германия).
Гаплогруппа H была обнаружена у представителей культуры линейно-ленточной керамики, живших ок. 7000 лет назад.
Субклада H2a1 обнаружена у представителя хвалынской культуры, жившего 6700 лет назад.
Субклада H2a3 обнаружена у представительниц анатолийского неолитического поселения Кумтепе, живших 6700 лет назад.
Субклада H2a1a определена у представителя среднестоговской культуры из Александрии (Alexandria) (Украина), жившего 6200 лет до настоящего времени.
H4 определена у обитательницы пещеры близ израильской деревни Пкиин, жившей ок. 4000 лет до нашей эры.
Субклады H2b, H6a1b, H13a1a1a обнаружены у представителей ямной культуры.
Субклада H2a была обнаружена у представителя культуры Ремеделло, жившего ок. 5300 лет назад.
Гаплогруппа H определена у представителей культуры воронковидных кубков.
Митохондриальная гаплогруппа H прослежена у обитателей верховий Западной Двины с 5120±120 лет назад (Сертея VIII) до VIII—X веков нашей эры (представитель культуры длинных курганов — H2).
H4a1 и H5b определены неолитических обитателей болгарской Дзулюницы (2800 лет до н. э.).
H2a выявлена у обитателя пещеры (Бадахшан, Афганистан), жившего ок. 4,5 тыс. лет назад.
Субклада H23 была обнаружена у представителя лужицкой культуры, жившего 1113—1021 лет до нашей эры.
H6a1a определена у представителя гальштатской культуры из Чехии (DA111, 2630 ± 48 лет до настоящего времени).
Субклады H5 и H6b определены у мумий из Абусира.
H4a1 определена у мумии из музея Ольстера (Белфаст, Северная Ирландия). Женщина с 33 зубами жила в Луксоре более 2600 лет назад (эпоха 25 династии).
H2 определена у мероитского образца MIS-TM и позднемероитского образца MIS-TMT из некрополя Миссиминия (Missiminia Necropolis) в районе Абри (Abri) в Верхней Нубии (350 год до н. э. — 350 год н. э.).
У славянских женских образца RISE568, RISE569 из Брандысек (район Кладно, Чехия), датированных VII—VIII веками нашей эры (660—770 calCE) митохондриальные гаплогруппы H и H1af2.
Гаплогруппа H обнаружена у 5 викингов с языческого захоронения Галгедил (Galgedil) на датском острове Фюн (700—1100 года).
Гаплогруппа H обнаружена у 73 % человек, похороненных в XIII–XVI веках в некрополе Сан-Мигель-де-Эреносар (San Miguel de Ereñozar) на северо-западе Испании.
Митохондрии в клетке
Митохондрия клетки печени крысы в разрезе.
Впервые митохондрии обнаружены в виде гранул в мышечных клетках в 1850 году.
Число митохондрий в клетке непостоянно. Их особенно много в клетках, у которых велика потребность в энергии. По своему строению митохондрии представляют собой органеллы, обычно сферической формы, встречающиеся в эукариотической клетке в количестве от нескольких сотен до 1—2 тысяч и занимающие 10—20 % её внутреннего объёма. Сильно варьируются также размеры (от 1 до 70 мкм) и форма митохондрий. В зависимости от того, в каких участках клетки в каждый конкретный момент времени происходит повышенное потребление энергии, митохондрии способны перемещаться по цитоплазме в зоны наибольшего энергопотребления, используя для движения структуры цитоскелета эукариотической клетки. В клетках растений и животных одновременно и примерно в равных количествах существуют три типа митохондриальных органелл: молодые протомитохондрии, зрелые митохондрии и старые постмитохондрии, деградирующие в липофусциновые гранулы.