Разница между днк и хромосомой
Содержание:
- Свойства гена
- Главное отличие — ДНК против хромосомы
- Механизмы транспорта ионов и молекул клеткой
- Прокариоты
- Клетки растений
- Рецепт ГМО: режь, исправляй, сшивай
- Всё началось с гороха
- Происхождение названия
- Ядро клетки
- Митоз и мейоз клетки
- Что такое плазмидная ДНК
- Снижаем риски: от ТРАНС к ЦИС и ниже
- Хромосомы человека
- Апоптоз
- Разница между хроматином и хромосомой
- Наследственность и хромосомы
Свойства гена
- дискретность — несмешиваемость генов;
- стабильность — способность сохранять структуру;
- лабильность — способность многократно мутировать;
- множественный аллелизм — многие гены существуют в популяции во множестве молекулярных форм;
- аллельность — в генотипе диплоидных организмов только две формы гена;
- специфичность — каждый ген кодирует свой признак;
- плейотропия — множественный эффект гена;
- экспрессивность — степень выраженности гена в признаке;
- пенетрантность — частота проявления гена в фенотипе;
- амплификация — увеличение количества копий гена[источник не указан 1498 дней].
Главное отличие — ДНК против хромосомы
Все известные живые организмы и многие вирусы используют ДНК в качестве своего генетического материала для хранения своей генетической информации. ДНК представляет собой двухвалентную структуру, которая существует в виде двойной спирали. Двойная спираль ДНК конденсируется с белками гистонов с образованием хромосомы. ключевое отличие между ДНК и хромосомой является то, что ДНК является неорганизованной структурой носителя генетической информации у большинства организмов, а хромосома — наиболее организованной структурой ДНК с гистонами в клетке. В дальнейшем, ДНК хранит генетические инструкции, тогда как хромосомы позволяют регулировать гены цепи ДНК.
Эта статья объясняет,
1. Что такое ДНК — определение, структура, особенности2. Что такое хромосома — определение, структура, особенности3. В чем разница между ДНК и хромосомой
Механизмы транспорта ионов и молекул клеткой
- Проницаемость мембраны клетки для ионов и молекул
- Транспортные каналы и белки клеточной мембраны
- Гидратная оболочка клетки и степень гидратации
- Мембранный потенциал и электрохимический градиент
- Калиевые каналы мембраны клетки
- Воротный механизм калиевых каналов — гейтинг
- Транспорт ионов натрия через плазматическую мембрану
- Эпителиальные натриевые каналы (ENaCs) мембраны клетки
- Кальциевые каналы мембраны клетки
- Хлоридные каналы мембраны клетки
- Транспорт воды через аквапорины мембраны клетки
- Формирование потенциала действия и потоки ионов
- Регуляция мышечного сокращения ионами кальция
- Перенос глюкозы через мембрану унипортным транспортом
- Механизм сопряженного транспорта через мембрану клетки
- Трансмембранный градиент ионов натрия и натрий зависимые переносчики
- Регуляция pH клетки и внеклеточной среды через обмен натрия
- Строение кальциевой АТФазы SERCA и механизм закачивания кальция
- Строение и функционирование натрий-калиевой АТФазы
- Строение и функционирование F1F0-АТФ-синтазы
- Строение и функционирование протонных насосов (H+-АТФазы, V-АТФазы)
- Современные возможности изучения ионных каналов
- Вывод и применение уравнения Нернста
- Механизмы входящего выпрямления калиевых каналов мембраны клетки
- Развитие муковисцидоза (кистозного фиброза) при мутации гена анионного канала CFTR
Прокариоты
- Схема строения прокариотической клетки
- Эволюция микроорганизмов — молекулярная филогенетика
- Требования к условиям жизни прокариот
- Сходства археев прокариот с эукариотами
- Строение гликокаликса — капсулы прокариот
- Строение и функции пептидогликана клеточной стенки бактерий
- Особенности клеточной оболочки грамположительных бактерий
- Особенности клеточной оболочки грамотрицательных бактерий
- Функции цитоплазматической мембраны бактерий (прокариот)
- Секреторные механизмы бактерий (прокариот)
- Строение пили, жгутиков бактерий и их участие в хемотаксисе
- Строение генома бактерий (плазмиды)
- Строение нуклеоида и цитоплазмы бактерий
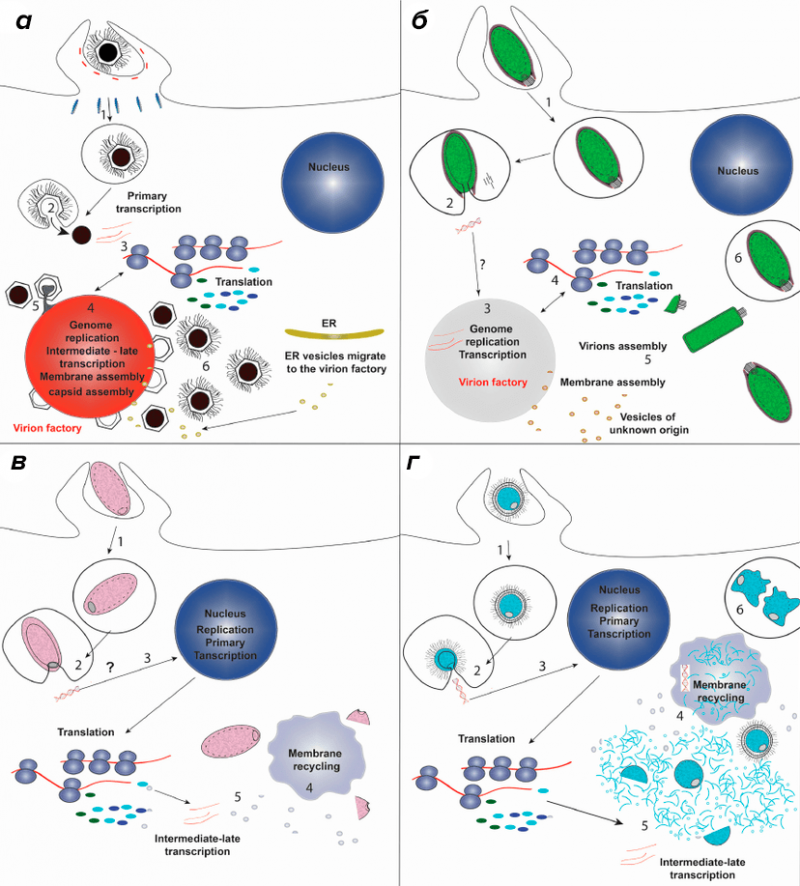
- Фабрики репликации ДНК бактерий
- Механизм сегрегации хромосом у бактерий (прокариот)
- Механизм деления бактерий (прокариот)
- Изменение развития бактерий (прокариот) в экстремальных условиях
- Изменение жизненного цикла развития Caulobacter crescentus
- Эндосимбиоз прокариот и эукариот
- Прокариоты как причины заболеваний у высших организмов
- Строение и образование биопленки микробов
- Перспективы изучения прокариот (бактерий)
Клетки растений
- Особенности строения клеток растений
- Как растут растения?
- Строение амикальных меристем и модулей роста растений
- Направление плоскости деления клеток растений и его значение
- Формирование направления плоскости деления клеток растений
- Механизм митоза клеток растений
- Механизм формирования новой клеточной стенки в ходе цитокенеза
- Механизм формирования клеточной пластинки в ходе цитокенеза
- Строение и функции плазмодесмы
- Механизм роста клеток растений набуханием вакуолей
- Целлюлозные микрофибриллы клеточной стенки клеток растений
- Изменение клеточной стенки клетки при ее росте
- Синтез целлюлозы в плазматической мембране клетки
- Строение и функции кортикальных микротрубочек
- Механизмы перестраивания кортикальных микротрубочек
- Механизм доставки материалов для роста плазматической мембраны клетки
- Механизм движения органелл и везикул клетки по актиновым путям
- Строение и функции ксилемы
- Точки роста и верхушечный рост растений
- Строение пластид и хлоропластов растений
- Строение хлоропластов и их функции
- Перспективы изучения клеток растений
Рецепт ГМО: режь, исправляй, сшивай
Так как же сделать ГМО из обычного растения? Рецепт довольно прост. Возьмите геном растительной клетки и добавьте в него «генетическую конструкцию» — последовательность ДНК, кодирующую производство нужного белка. Доставить ген можно с помощью вектора — молекулы ДНК или РНК, способной «размножаться» и переносить чужеродный наследственный материал из клетки в клетку. Таким образом можно добавить сразу несколько генов, например, с помощью вектора на основе кольцевой бактериальной плазмиды.
На первый взгляд, все просто, если не задумываться о том, как вставить новый генный фрагмент именно в тот участок ДНК растительной клетки, который нам нужен. А ведь в этом и заключается самая сложная задача редактирования генома, результатом которого являются современные ГМО.
Для того, чтобы расщеплять молекулы ДНК в точно заданных участках, сначала стали использовать рестриктазы — ферменты-«ножницы», способные узнавать определенные последовательности нуклеотидов (строительных блоков ДНК). Функцию сшивки ДНК-цепи исполняли другие ферменты — ДНК-лигазы, призванные исправлять (репарировать) повреждения в структуре ДНК.
Сегодня, как и 30–40 лет назад, эти методы активно используют для получения новых вариантов бактериальных и вирусных геномов. А вот для успешной работы с геномами высших организмов (таких как растения, животные и мы с вами) этих инструментов оказалось недостаточно. Дело в том, что рестриктазы способны узнавать лишь короткие последовательности ДНК, что вполне достаточно для эффективного расщепления коротких ДНК-цепей бактерий, где такие участки встречаются нечасто. Но геномы высших организмов содержат огромное множество коротких последовательностей нуклеотидов, узнаваемых рестриктазами, поэтому «прицельность» метода оказывается очень плохой.
Для редактирования таких геномов пришлось создавать свои инструменты точечного воздействия на ДНК: сначала олигонуклеотид-направленный мутагенез растений, затем сайт-направленный мутагенез с использованием ферментов-нуклеаз с «цинковыми пальцами», TALENs-нуклеаз и даже мегануклеаз (Закиян, 2014; Daboussi, 2015). Но лишь с открытием в 2012–2013 гг. знаменитой технологии CRISPR/Cas9 ученые вплотную подошли к точному исправлению или редактированию генов и геномов (Cong, 2013). Возможность вносить контролируемые изменения в наследственную информацию живых клеток стала настоящим прорывом и повлекла за собой глобальные изменения в селекции.
Всё началось с гороха
Аббат Грегор Мендель, австрийский ботаник и биолог, заметил, что потомство не всегда повторяет признаки, которыми обладали родители. Чтобы понять взаимосвязь, Мендель стал выращивать горох, скрещивать различные растения и отслеживать частоту наследования признаков.
Мендель доказал, что отдельные признаки (цвет, форма цветка и т.д.) могут наследоваться независимо. Он вывел теорию доминантных и рецессивных признаков, описал явление прерывистого наследования, математически интерпретировал результаты своих экспериментов.
Труды Менделя впервые опубликовали в 1866 году. Именно его считают основоположником генетики.
До этого ученые считали, что родительские признаки смешиваются подобно жидкости и потомки наследуют именно такой «коктейль». Теория пангенезиса, которую Чарльз Дарвин сформулировал в 1868 году, также следует этой концепции.
Впрочем, Дарвин считал, что «коктейль» состоит из мельчайших отдельных частиц – геммул. Они смешиваются во время зачатия. В целом ученый был недалек от истины.
Собственно термин «ген» в 1909 году ввел Вильгельм Йоханнсен. До этого признаки называли пангенами.
Происхождение названия
Термин «геном» был предложен Гансом Винклером в 1920 году в работе, посвящённой межвидовым амфидиплоидным растительным гибридам, для описания совокупности генов, заключённых в гаплоидном наборе хромосом организмов одного биологического вида. В Оксфордском энциклопедическом словаре указано, что термин образован слиянием слов «ген» и «хромосома». Однако Джошуа Ледерберг и Алекса T. МакКрэй считают, что ботаник Г. Винклер должен был быть знаком с ботаническими терминами «ризом», «таллом», «трахеом» и т. д. Все эти термины возникли до 20-х годов XX века, и суффикс «-ом» в них означает объединение частей в целое, например, «ризом» означает всю корневую систему растения. Таким образом, «геном» можно понимать как объединение генов в целое.
До недавнего времени термин «геном» использовался в двух смыслах. У эукариот геном соответствовал гаплоидному набору хромосом с локализованными в них генами. В генетике бактерий и вирусов термин «геном» употреблялся для обозначения совокупности наследственных факторов одной хромосомы или группы сцепления прокариот. Позже в генетике бактерий семантика термина «геном» претерпела дрейф в сторону обозначения всей наследственной конституции клетки, включая самые разные внехромосомные факультативные элементы. Постепенно в этом смысле термин «геном» стали употреблять и в генетике эукариот.
Первоначальный смысл этого термина указывал на то, что понятие генома, в отличие от генотипа, является генетической характеристикой вида в целом, а не отдельной особи. С развитием молекулярной генетики значение данного термина изменилось. В настоящее время под «геномом» понимают совокупность наследственного материала отдельного представителя вида, примером может служить международный проект 1000 геномов, целью которого является секвенирование геномов 1000 человек.
Ядро клетки
- Строение ядра клетки и его функции
- Факторы влияющие на размер и форму ядра клетки
- Локализация хромосом в ядре клетки
- Субкомпартменты ядра — ядрышки
- Функции ядерных сайтов и ядерного матрикса
- Строение и функции ядерной оболочки
- Строение и функции ядерной ламины
- Транспорт между ядром и цитоплазмой через ядерные поровые комплексы (ЯПК)
- Строение комплекса ядерной поры (ЯПК, ядерного порового комплекса)
- Нуклеопорины ядерных поровых комплексов (ЯПК)
- Механизм транспорта белков в ядро через ядерные поры
- Функции сигнала ядерной локализации (NLS)
- Механизм импорта белков в ядро через рецепторы NLS
- Механизм экспорта белка из ядра клетки
- Механизм контроля ядерного транспорта Ran-ГТФазой
- Модели механизма ядерного транспорта
- Механизм регулирования ядерного транспорта
- Механизм экспорта РНК из ядра клетки
- Сборка и экспорт рибосом
- Механизм экспорта тРНК из ядра
- Механизм экспорта иРНК из ядра
- Механизм транспорта гетерогенных ядерных рибонуклеопротеиновых частиц (гяРНП)
- Факторы участвующие в экспорте иРНК
- Малые ядерные РНК (UмяРНК) и малые ядерные рибонуклеопротеидные частицы (мяРНП)
- Механизм транспорта микроРНК
- Перспективы изучения ядра клетки
- Значение генетики в медицине. Сферы применения
- Значение современной генетики в медицине. Задачи
- Геном человека. Особенности
- Строение ДНК человека. Особенности структуры
- Строение хромосом человека. Особенности организации
- Организация генома человека. Особенности
- Кариотип человека. Особенности набора хромосом
- Информационное содержание генома человека. Особенности генов
- Структура генов человека. Организация
- Семейства генов человека. Особенности
- Псевдогены человека. Особенности
- Экспрессия генов: транскрипция, трансляция, процессинг
- Механизмы экспрессии гена бета-глобина. Этапы синтеза b-глобина
- Генетика иммуноглобулинов и рецепторов Т-клеток. Особенности экспрессии генов
- Термины геномики и молекулярной генетики. Определения
- Анализ ДНК и РНК. Молекулярное клонирование
а) Хромосомы ядра клетки
- Строение хромосом и их размеры
- Строение хромосом: хроматин, центромера, теломера
- Строение хроматина и его формы
- G-полосы хромосом и их значение
- Петли и домены ДНК для ее спирализации
- Механизм соединения ДНК с ядерным матриксом при помощи MAR (matrix attachment regions)
- Локализация и функции центромер хромосом
- Функции ДНК центромер Saccharomyces cerevisiae
- Белки связанныые с центромерами
- Повторы ДНК в области центромеров
- Функция и репликация теломер хромосом
- Замыкание концов хромосом теломерами
- Хромосомы типа ламповых щеток
- Строение политенных хромосом
- Значение пуфов (вздутий) политенных хромосом
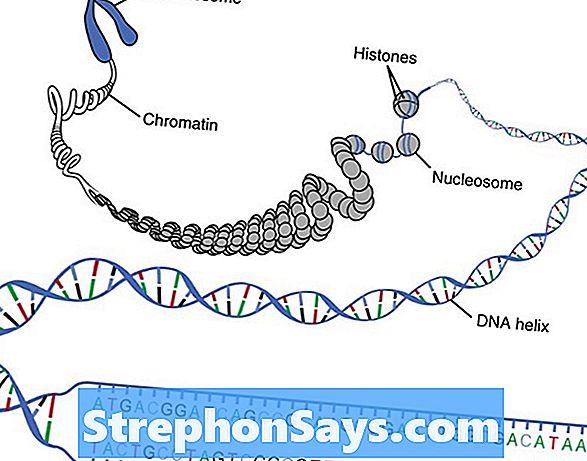
- Нуклеосома как структурная единица хромосомы
- Содержание ДНК в нуклеосоме
- Коровая и линкерная ДНК нуклеосом
- Структура ДНК нуклеосом
- Строение октамера гистонов нуклеосомы
- Строение фибрилл хроматина и их нуклеосомы
- Сборка нуклеосом при репликации ДНК и хроматина
- Позиционирование нуклеосом на ДНК
- Структура домена ДНК с активными генами
- Все ли гены организованы в нуклеосомы?
- Механизм удаления гистонов с нуклеосом при транскрипции
- Факторы удаления и сборки гистонов нуклеосом
- Сайты ДНК с повышенной чувствительностью к ДНКазе (нуклеазе)
- Механизм ремоделирования хроматина
- Механизм ацетилирования гистонов
- Строение и образование гетерохроматина
- Взаимодействие белков с гистонами при образовании гетерохроматина
- Механизмы инактивирования Х-хромосомы
- Значение белков SMC (конденсинов, когезинов) в структуре хромосомы
- Перспективы изучения хромосом
Митоз и мейоз клетки
- Схема митоза и его фазы
- Характеристика фаз митоза
- Видео митоза клетки и его стадий
- Образование и строение веретена деления
- Микротрубочки веретена деления
- Образование и строение центросомы
- Механизмы формирования митотического веретена деления
- Механизмы стабилизации веретена деления
- Значение центромеры и кинетохора
- Образование и строение кинетохора
- Захват и стабилизация микротрубочек кинетохорами
- Типы прикрепления хромосом к кинетохорам
- Механизмы сокращения и удлинения нитей кинетохора
- Механизмы движения хромосом к полюсу
- Механизм конгрессии хромосом
- Регуляция конгрессии хромосом
- Контроль перехода метафазы в анафазу кинетохорами
- Характеристика фаз анафазы митоза
- Что происходит в телофазе митоза?
- Механизмы и этапы цитокенеза
- Механизмы образования сократимого кольца
- Механизм деления клетки сократимым кольцом
- Распределение органелл при делении клеток в ходе митоза
- Перспективы изучения митоза
- Клеточный цикл — митоз: описание фаз G0, G1, G2, S
- Гаметогенез — мейоз: фазы, особенности у мужчин, женщин
- Митотический цикл клетки. Фазы клеточного цикла
- Фазы митоза клетки. Особенности
- Фазы мейоза половых клеток. Особенности
- Гаметогенез: сперматогенез, овогенез, оплодотворение
- Медицинское и биологическое значение митоза и мейоза
Что такое плазмидная ДНК
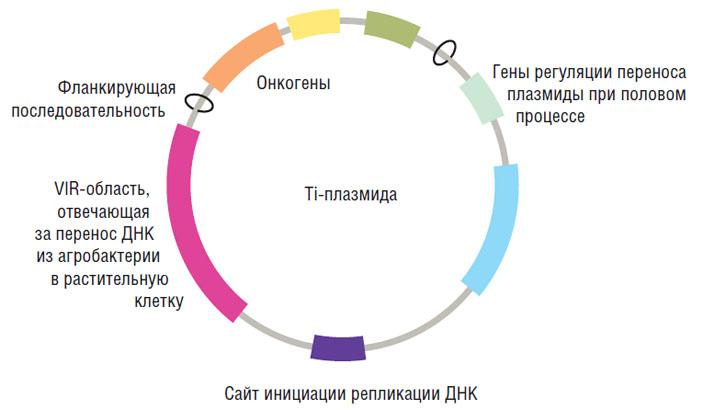
Плазмидная ДНК — это тип ДНК, отделенный от геномной ДНК. Следовательно, это форма внехромосомной ДНК. Оно всегда круглое и естественно встречается внутри прокариотических клеток. Кроме того, плазмидная ДНК является небольшой молекулой по сравнению с хромосомами. Конкретная клетка может содержать переменное число плазмид определенного типа. Как правило, плазмидная ДНК является самореплицирующейся ДНК, поскольку она содержит источник репликации. Следовательно, плазмиды могут реплицироваться независимо от геномной ДНК.

Рисунок 1: Плазмидная ДНК
Плазмидная ДНК кодирует несколько генов, чьи генные продукты не являются необходимыми для общего функционирования клетки. Эти гены кодируют устойчивость к антибиотикам, устойчивость к металлам, фиксацию азота и выработку токсинов. Это означает, что клетка требует только эти генные продукты для выживания при определенных условиях. Как прокариотические, так и эукариотические клетки поглощают плазмидную ДНК в естественных или искусственно индуцированных условиях. Следовательно, трансформация рекомбинантных плазмид в организм хозяина может ввести новые гены в организм в процессе, называемом генной инженерией.
Снижаем риски: от ТРАНС к ЦИС и ниже
Сейчас прохождение всех тестов на биобезопасность и вывод на рынок генетически модифицированных организмов, в том числе растений, жестко регулируется на международном уровне. В этом вопросе правовая база ЕС опирается на директиву Европейского парламента и Совета Европейского союза от 12 марта 2001 г. «О преднамеренном выпуске в окружающую среду генетически модифицированных организмов». Примечательно, что данный нормативный документ исключает из перечня ГМО организмы, полученные путем скрещивания, экстракорпорального оплодотворения, полиплоидной индукции, возникновения мутаций и слияния протопластов скрещиваемых видов (соматическая гибридизация).
Законодательство РФ в области ГМО растительного происхождения регулируют 4 федеральных закона и 6 постановлений Правительства РФ, в том числе федеральный закон № 86-ФЗ «О государственном регулировании в области генно-инженерной деятельности» от 5 июля 1996 г. Ожидается вступление в силу постановления, позволяющего узаконить возделывание ГМ-культур на территории России, которые пока можно выращивать только на опытных участках. Для ввоза в нашу страну сегодня разрешены 22 линии пищевых и кормовых ГМ-растений, среди которых кукуруза, картофель, соя, сахарная свекла и рис, а любые ГМО и ГМ-продукты должны проходить обязательную регистрацию.
В свою очередь, мировое ученое сообщество считает, что нужно различать ГМО по способу получения и делать послабления для продуктов, полученных умеренным вмешательством. Так появилась система деления ГМО на три вида: ТРАНС, ЦИС и ИНТРА.
Трансгенными сегодня называют организмы с искусственно введенными генами, которые в принципе не могут быть приобретены путем естественного скрещивания. Это могут быть гены растений других видов или животных, например рис, в геном которого встроен ген кукурузы. Потенциальная опасность трансгенных культур в том, что приобретенные таким образом новые качества могут повлиять на пригодность к использованию в пищевых или кормовых целях, а затем передаться диким родственникам, что может иметь непредсказуемые последствия для природных экосистем
По этой причине законодательные и регулирующие органы развитых стран уделяют большое внимание биобезопасности таких культур, чтобы снизить риск экологических сдвигов
В геном цисгенных растений могут быть введены гены организмов того же или близких видов, с которыми возможно скрещивание в естественных условиях. При этом сам целевой ген не должен быть видоизменен или оторван от своих регуляторных последовательностей. Пример цисгенного растения — картофель, не подверженный картофельной гнили благодаря встраиванию генов диких видов картофеля из Анд, устойчивых к этому заболеванию. Такой картофель сейчас создается в Бельгии (VIB’s fact series, 2015)
Важно, что цисгенезис не привносит в организм растения принципиально новых для него признаков и, по сути, аналогичен традиционному скрещиванию с родственными дикими формами
Интрагенезис можно считать продолжением концепции цисгенезиса, но в этом случае в ДНК растения встраивают его собственный ген, совмещенный с регуляторными участками других его генов. В ходе такой модификации искусственно создаются новые комбинации из уже имеющихся в растении участков ДНК (Holme, 2013). Подобное изменение регуляции активности генов позволяет усиливать полезные признаки (например, способность накапливать витамины в листьях) или, напротив, устранять или сводить к минимуму нежелательные.
Между тем при современном регулировании оборота ГМО-различия между трансгенными и цисгенными растениями не учитываются, хотя эти типы кардинально различаются. Из-за жестких рамок, установленных законодательством, получение и использование цисгенных растений серьезно затруднено, что может заблокировать или значительно отсрочить проведение дальнейших исследований по улучшению сортов сельскохозяйственных культур. Пока лишь в Канаде контроль за цисгенными растениями менее строг по сравнению с трансгенными (Schouten, 2006).
Хромосомы человека
Нормальный кариотип человека представлен 46 хромосомами. Это 22 пары аутосом и одна пара половых хромосом (XY в мужском кариотипе и XX — в женском). В приведённой ниже таблице показано число генов и оснований в хромосомах человека.
Изображение 46 (23 пар) хромосом женского кариотипа человека, полученное с помощью FISH с флуоресцентно-мечеными Alu-повторами. Alu-повторы показаны зелёным цветом, ДНК — красным. У человека самая длинная 1-я хромосома примерно в 5 раз длиннее самой короткой 21-й хромосомы.
| Хромосома | Всего оснований | Количество генов | Количество белок-кодирующих генов |
|---|---|---|---|
| 249250621 | 3511 | 2076 | |
| 243199373 | 2368 | 1329 | |
| 198022430 | 1926 | 1077 | |
| 191154276 | 1444 | 767 | |
| 180915260 | 1633 | 896 | |
| 171115067 | 2057 | 1051 | |
| 159138663 | 1882 | 979 | |
| 146364022 | 1315 | 702 | |
| 141213431 | 1534 | 823 | |
| 135534747 | 1391 | 774 | |
| 135006516 | 2168 | 1914 | |
| 133851895 | 1714 | 1068 | |
| 115169878 | 720 | 331 | |
| 107349540 | 1532 | 862 | |
| 102531392 | 1249 | 615 | |
| 90354753 | 1326 | 883 | |
| 81195210 | 1773 | 1209 | |
| 78077248 | 557 | 289 | |
| 59128983 | 2066 | 1492 | |
| 63025520 | 891 | 561 | |
| 48129895 | 450 | 246 | |
| 51304566 | 855 | 507 | |
| X-хромосома | 155270560 | 1672 | 837 |
| Y-хромосома | 59373566 | 429 | 76 |
Апоптоз
- В чем отличие апоптоза от некроза? Причины апоптоза
- Типы каспаз и их участие в регуляции апоптоза
- Механизмы активации каспаз апоптоза
- Функции белкового ингибитора апоптоза (IAP)
- Каспазы участвующие в воспалении и обмене цитокинов
- Пути апоптоза и их схемы
- Механизм апоптоза вызванного рецептором TNFR1
- Механизм и схема митохондриального пути апоптоза
- Механизм повышения проницаемости наружной мембраны митохондрий (МОМР)
- Механизм активации Bax и Bak белками семейства Bcl-2
- Механизм активации каспазы цитохромом с апоптосомы
- Механизм ингибирования белками Smac и Omi белковых ингибиторов апоптоза IAPs
- Связующая функция белка Bid между путями апоптоза
- Механизм гибели клетки при повышении проницаемости наружной мембраны митохондрий (МОМР)
- Разрыв наружной мембраны митохондрий как причина апоптоза
- Механизм апоптоза у нематод С. elegans
- Отличия апоптоза насекомых от апоптоза млекопитающих
- Механизм удаления погибающих (апоптических) клеток
- Значение апоптоза в развитии вирусных инфекций и рака
- Влияние погибающих (апоптических) клеток на иммунитет
- Перспективы изучения апоптоза
Разница между хроматином и хромосомой
Определение
хроматина: Молекулы ДНК в геноме упакованы гистонами, образующими хроматин.
Хромосома: Высшая упакованная структура ДНК появляется в метафазе клеточного деления.
период
хроматина: Хроматин появляется в интерфазе клеточного цикла.
Хромосома: Хромосомы появляются во время метафазы и существуют в анафазе ядерного деления.
хроматина: Хроматин состоит из нуклеосом.
Хромосома: Хромосомы конденсируются в хроматиновые волокна.
Condensity
хроматина: Хроматин конденсируется в 50 раз по сравнению с нормальной двойной спиралью ДНК.
Хромосома: Хромосомы сгущаются в 10000 раз больше, чем обычная двойная спираль ДНК.
Внешность
хроматина: Хроматиновые волокна представляют собой тонкие, длинные, незакрученные структуры.
Хромосома: Хромосомы представляют собой толстые, компактные, лентообразные структуры.
хроматина: Хроматин представляет собой одиночные непарные волокна.
Хромосома: Хромосома существует в виде пары.
функция
хроматина: Хроматин позволяет упаковывать генетический материал в ядро, регулируя экспрессию генов.
Хромосома: Хромосомы обеспечивают правильное расположение генетического материала в клеточном экваторе, чтобы обеспечить равное разделение генетического материала между двумя клетками.
Метаболическая активность
хроматина: Хроматин обеспечивает репликацию ДНК, экспрессию генов и рекомбинацию.
Хромосома: Хромосомы не проявляют метаболической активности.
подтверждение
хроматина: Хроматин состоит из двух подтверждений: эухроматин и гетерохроматин.
Хромосома: Хромосома обычно гетерохроматическая. Он состоит из таких форм, как метацентрическая, субметацентрическая, акроцентрическая и телоцентрическая.
Визуализация
хроматина: Хроматин можно наблюдать под электронным микроскопом в виде структуры бус и струн.
Хромосома: Хромосому можно наблюдать под световым микроскопом в ее классической структуре из четырех плеч.
Заключение
Хроматин и хромосома — это два типа конденсированных структур молекул ДНК. Интерфазная ДНК существует в виде нитевидной структуры, известной как хроматин. Основная цель хроматина заключается в упаковке двухцепочечной ДНК в ядро клетки с помощью гистоновых белков. Молекулы ДНК конденсированы в 50 раз по сравнению с нормальной структурой в волокнах хроматина. Хромосома появляется только в метафазе клеточного цикла, достигая своей максимальной конденсированной структуры. Основная цель хромосомы состоит в том, чтобы обеспечить разделение двойного генетического материала между двумя дочерними клетками. Основное различие между хроматином и хромосомой заключается в их конденсации и функции во время клеточного цикла.
Ссылка:1. «Хроматин». Википедия. Н.П .: Фонд Викимедиа, 27 февраля 2017 г. Веб. 6 марта 2017 г. 2. «Хромосома». Википедия. Н.П .: Фонд Викимедиа, 3 марта 2017 г. Веб. 6 марта 2017 г.
Изображение предоставлено:1. «Нуклеосома 1KX5 2» Зефириса из Википедии на английском языке
Наследственность и хромосомы
Хранителями наследственной информации являются хромосомы – микроскопические тельца в ядре клетки (рис.1). В каждой клетке любого организма конкретного вида содержится определенное число хромосом. У человека их 46. Поскольку хромосомы всегда парны (имеется по две хромосомы каждого сорта), то можно сказать, что у человека 23 пары хромосом. Подавляющее большинство клеток человеческого организма содержат все 46 хромосом (диплоидный набор), и только половые клетки – гаметы – имеют по 23 хромосомы (гаплоидный набор). При оплодотворении мужская и женская половые клетки сливаются, поэтому оплодотворенная яйцеклетка, как и все клетки развивающегося из него нового организма, получают 46 хромосом, половину – от матери, половину – от отца.
Хромосома – это гигантская молекула дезоксирибонуклеиновой кислоты (ДНК), скрученная особым образом. На отдельных участках молекулы ДНК (соответственно, и хромосомы) закодирована наследственная информация. Участок молекулы ДНК, в котором зашифрована единица наследственной информации, называется геном (рис. 2). Один ген хранит информацию о строении определенной молекулы белка, входящей в состав тела человека. Совокупность генов, то есть вся информация, заключенная в хромосомах, получила название геном.