Микротрубочки
Содержание:
- Микротрубочки, их строение и функции.
- У растений
- Строение и свойства микротрубочек
- Какие функции выполняют микротрубочки?
- Примечания[ | код]
- Строение
- Белки микротрубочек
- Микротрубочки стареют?
- Рибосомы
- Функция
- Чего пока не может эксперимент и как помогает теория?
- У животных
- Взаимодействие актин-миозиновой и тубулин-динеиновой комплекса
- Динамическая нестабильность[ | код]
- Микрофиламенты
- Расположение микротрубочек
Микротрубочки, их строение и функции.
Проведенный в последнее время детальный сравнительно-цитологический анализ организации жгутиков сперматозоидов у разных многоклеточных животных показал возможность существенных изменений стандартной формулы 9 + 2 даже у близкородственных животных.
В жгутиках сперматозоидов некоторых групп животных две центральные микротрубочки могут отсутствовать, а их роль выполняют цилиндры из электронно-плотного вещества. Среди низших многоклеточных (турбеллярии и близкие к ним группы) подобного рода модификации распределены у отдельных видов животных мозаично и, вероятно, полифилетичны по своему происхождению, хотя у всех этих видов образуются сходные морфологические структуры.
Еще более значительные модификации постоянных тубулин-динеиновых систем наблюдаются в щупальцах некоторых простейших. Здесь эта система представлена группой антипараллельных микротрубочек. Динеиновые структуры, связывающие микротрубочки, имеют отличный от динеиновых «рук» ресничек и жгутиков характер расположения, хотя принцип работы динеин-тубулиновой системы ресничек, жгутиков и щупалец простейших, по-видимому, сходен.
У растений
Высшие растения не имеют центриолей. Сайты нуклеации микротрубочек, содержащие γ-тубулин, на разных стадиях клеточного цикла могут быть диффузно распределены в цитоплазме или заякорены на различных внутренних мембранах клетки: на эндоплазматической сети и на ядерной мембране. Кроме того, в телофазе ЦОМТ сосредотачиваются в районе фрагмопласта. У печёночного мха Dumortiera hirsuta в клетках перед началом мейоза диффузные ЦОМТ сосредотачиваются на поверхности единственной пластиды, а после окончания деления пластиды перемещаются в цитоплазму, где формируют веретено деления; затем, в телофазе I, ЦОМТ располагаются на поверхности ядер и в районе фрагмопласта.
Строение и свойства микротрубочек
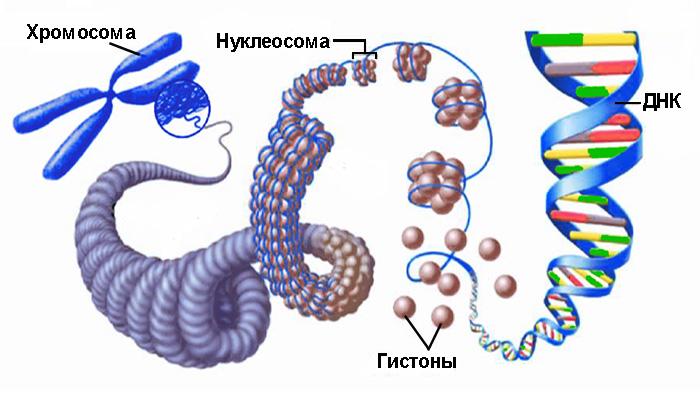
Микротрубочки — это линейные полимеры. Они построены из димеров белка тубулина, которые образуют 13 цепей — протофиламентов (рис. 1). Каждый из них по бокам связан с двумя другими, и вся конструкция замкнута в цилиндр диаметром 25 нм. Такое строение обеспечивает микротрубочке прочность и большую изгибную жесткость: она может оставаться почти абсолютно прямой в масштабе клетки. Чтобы представить, насколько микротрубочка сложно сгибаема, мысленно увеличим ее до размеров стержня диаметром спагетти (около 2 мм). Такая «спица» не прогибалась бы, будь она длиной даже в сотни метров (высота современных небоскребов)! Жесткость позволяет микротрубочкам выполнять роль длинных прямых направляющих, которые организуют движение органелл внутри клетки. Остальные элементы цитоскелета (актиновые и промежуточные филаменты) существенно более гибкие, поэтому, как правило, используются клеткой в других целях.
Димер тубулина, из которого строится микротрубочка, состоит из мономеров двух типов. Внутри каждого протофиламента α-мономеры одного димера соединяются с β-мономерами соседнего. Поэтому по всей длине микротрубочки, содержащей десятки и сотни тысяч димеров тубулина, все они ориентированы одинаково. Тот конец микротрубочки, к которому обращены α-тубулины, называется минус-концом, а противоположный — плюс-концом. Благодаря такому упорядоченному расположению димеров микротрубочка имеет полярность, что обеспечивает направленность транспорта. Моторные белки, которые участвуют в перемещении «грузов» из одной части клетки в другую, «шагают» по микротрубочке, перетаскивая свою «ношу» за собой, как правило, только в одном направлении. Например, белок динеин двигает органеллы к минус-концу микротрубочки, а кинезин — к плюс-концу. Часто микротрубочки расположены в клетке радиально, а их плюс-концы направлены к ее периферии. Таким образом, кинезины осуществляют транспортировку из центра к внешней мембране, а динеины — от нее внутрь клетки. Поразительно, но в отростках аксонов везикулы и органеллы могут направленно передвигаться по микротрубочкам на расстояния в сотни микрометров и больше.
Какие функции выполняют микротрубочки?
Для них характерны самосборка и саморазборка, причем разборка происходит до тубулиновых димеров. Соответственно микротрубочки могут быть представлены большим или меньшим количеством в связи с преобладанием процессов либо саморазборки, либо самосборки микротрубочек из фонда глобулярного тубулина гиалоплазмы.
Интенсивные процессы самосборки микротрубочек обычно приурочены к местам крепления клеток к субстрату, т. е. к местам усиленной полимеризации фибриллярного актина из глобулярного актина гиалоплазмы.
Такая корреляция степени развития этих двух механохимических систем не случайна и отражает их глубокую функциональную взаимосвязь в целостной опорно-сократимой и транспортной системе клетки.
Материал с сайта http://Wiki-Med.com
На этой странице материал по темам:
В эту группу органоидов входят рибосомы, микротрубочки и микрофиламенты, клеточный центр.
Рибосома
Рибосомы (рис. 1) присутствуют в клетках как эукариот, так и прокариот, поскольку выполняют важную функцию в биосинтезе белков.
В каждой клетке имеются десятки, сотни тысяч (до нескольких миллионов) этих мелких округлых органоидов. Это округлая рибонуклеопротеиновая частица. Диаметр ее составляет 20—30 нм. Состоит рибосома из большой и малой субъединиц, которые объединяются в присутствии нити м-РНК (матричной, или информационной, РНК). Комплекс из группы рибосом, объединенных одной молекулой м-РНК наподобие нитки бус, называется полисомой. Эти структуры либо свободно расположены в цитоплазме, либо прикреплены к мембранам гранулярной ЭПС (в обоих случаях на них активно протекает синтез белка).

Рис.1.Схема строения рибосомы, сидяшей на мембране эндоплазматической сети: 1 — малая субъединииа; 2 иРНК; 3 — аминоацил-тРНК; 4 — аминокислота; 5 — большая субъединица; 6 — — мембрана эндоплазматической сети; 7 — синтезируемая полипептидная цепь
Полисомы гранулярной ЭПС образуют белки, выводимые из клетки и используемые для нужд всего организма (например, пищеварительные ферменты, белки женского грудного молока).
Кроме этого, рибосомы присутствуют на внутренней поверхности мембран митохондрий, где также принимают активное участие в синтезе белковых молекул.
Примечания[ | код]
- ↑ Shaw et al., 2003.
- Weber and Westermann, 2003.
- Ueda, 1999.
- Транзиентная экспрессия (от лат. transitus — переход, прохождение и expressio — выражение) — непродолжительная (временная) экспрессия клонированного гена, введенного в непермиссивную клеточную систему с помощью трансфекции и не интегрированного с геномом клетки-хозяина.
- Marc et al., 1998.
- ↑ Wasteneys and Yang, 2004.
- Barlow and Balushka, 2000.
- ↑ Goddard et al., 1994.
- Lloyd, 1994.
- Sugimoto et al., 2000.
- ↑ Dolan et al., 1993.
- Рейвн и др., 1990.
- Dixit et al., 2006.
- ↑ Yuan et al., 1994.
- Dixit and Cyr , 2004.
- ↑ Lloyd., 1994.
- Baskin et al., 2004.
- Burk et al., 2006.
- ↑ Dolan et al., 1994.
- Sieberer et al., 2002.
- ↑ Van Bruaene et al., 2004.
- Sieberer et al., 2005
- Balusˇka et al., 2000.
- Geitmann and Emons, 2000.
- Bibikova et al., 1999.
- ↑ Vassileva et al., 2005.
Строение
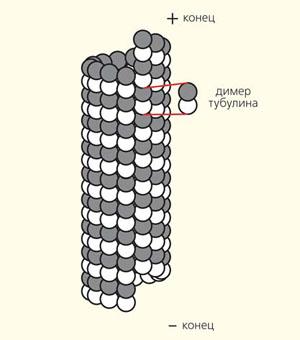
Микротрубочки — структуры, в которых 13 тубулиновых α- и β-гетеродимера заключены по кругу полого цилиндра. Внешний диаметр цилиндра составляет около 25 нм, внутренний — около 15 нм. Один из концов микротрубочки, называется положительным концом (или плюс концов), постоянно присоединяет к себе свободный тубулин. От другого конца — отрицательного (минус-конца) — тубулина субъединицы отщепляются.
В образовании микротрубочки in vitro выделяют три фазы:
- замедленная фаза, или нуклеация. Это этап зарождения микротрубочки, когда молекулы тубулина начинают соединяться в более крупные образования. Такое соединение происходит медленнее, чем присоединение тубулина к уже собранной микротрубочки, поэтому фаза и называется замедленной.
- фаза полимеризации, или элонгация. Если концентрация свободного тубулина высока, его полимеризация происходит быстрее, чем деполимеризацию на негативном конце, за счет чего микротрубочки увеличивается. По мере ее роста концентрация тубулина падает до критической и скорость роста замедляется до вступления в следующей фазе;
- фаза стабильного состояния. Деполимеризацию уравновешивает полимеризацию, и рост микротрубочки останавливается. Лабораторные исследования показывают, что сборка микротрубочек из тубулина происходит только в присутствии гуанозинтрифосфат и ионов магния, оптимально при температуре 37 ° C.
Белки микротрубочек
Система микротрубочек является вторым компонентом опорно-сократимого аппарата, находящаяся, как правило, в тесном контакте с микрофибриллярным компонентом. Стенки микротрубочек образованы в поперечнике чаще всего 13 димерными глобулами белка, каждая глобула состоит из α- и β-тубулинов (рис. 6). Последние в большинстве микротрубочек расположены в шахматном порядке. Тубулин составляет 80% белков содержащихся в микротрубочках. Остальные 20% приходятся на долю высокомолекулярных белков МАР1, МАР2 и низкомолекулярного тау-фактора. МАР-белки (microtubule-associated proteins- белки, связанные с микротрубочками) и тау-фактор представляют собой компоненты, необходимые для полимеризации тубулина. В их отсутствие самосборка микротрубочек путем полимеризации тубулина крайне затруднена и образующиеся микротрубочки сильно отличаются от нативных.
Микротрубочки — очень лабильная структура, так, микротрубочки теплокровных животных, как правило, разрушаются на холоде. Существуют и холодоустойчивые микротрубочки, например в нейронахцентральной нервной системы позвоночных их количество варьирует от 40 до 60%. Термостабильные и термолабильные микротрубочки не различаются по свойствам входящего в их состав тубулина; по-видимому, эти отличия определяются добавочными белками. В нативных клетках по сравнению с микрофибриллами основная часть микротрубочковой субмембранной системы располагается в более глубоко лежащих участках цитоплазмы Материал с сайта http://wiki-med.com
Микротрубочки стареют?
Модель ГТФ-«шапочки» привлекла внимание исследователей прежде всего потому, что позволила объяснить, почему микротрубочка может устойчиво расти и укорачиваться и почему между этими фазами возможны переходы — катастрофы и спасения. В 1995 г
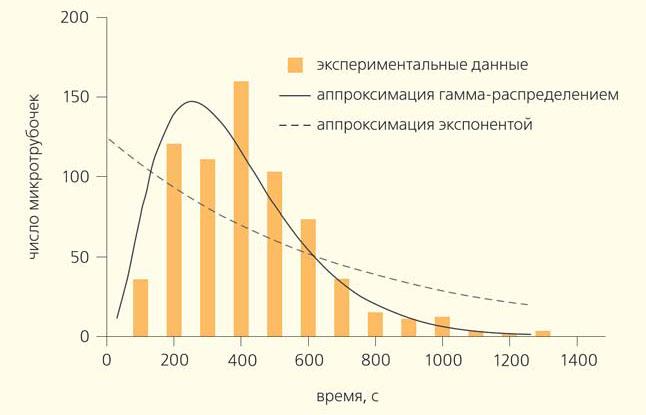
Д. Одде (D. Odde) с соавторами провел простой, но важный эксперимент []. Они наблюдали за ростом микротрубочек в пробирке и решили построить распределение их длин. Оно предполагалось экспоненциальным, но оказалось, что у него есть пик (рис. 3). Значит, в начале роста микротрубочки имеют очень маленькую вероятность испытать катастрофу, а дальше, по мере их роста, эта вероятность повышается. Если пересчитать распределение длин микротрубочек в частоты катастроф, то получится возрастающая зависимость частоты катастроф от времени. Этот эффект назвали «старением» микротрубочек — они как будто «портятся» со временем. Иначе говоря, «молодые» микротрубочки могут расти стабильно, а «старые» уже более склонны к разборке. Необычное распределение времен жизни микротрубочек хорошо аппроксимируется гамма-распределением, которое характеризует процессы с фиксированным количеством последовательных шагов. Поэтому возникла идея, что лучше всего результаты проведенного эксперимента описывает теория, согласно которой катастрофа микротрубочки происходит за три последовательных стадии, когда в ней накопились определенные дефекты неизвестной природы []. Эта гипотеза, исходно достаточно сомнительная, тем не менее существенно подогрела интерес к исследованию динамики микротрубочек на уровне отдельных димеров тубулина
В 1995 г. Д. Одде (D. Odde) с соавторами провел простой, но важный эксперимент []. Они наблюдали за ростом микротрубочек в пробирке и решили построить распределение их длин. Оно предполагалось экспоненциальным, но оказалось, что у него есть пик (рис. 3). Значит, в начале роста микротрубочки имеют очень маленькую вероятность испытать катастрофу, а дальше, по мере их роста, эта вероятность повышается. Если пересчитать распределение длин микротрубочек в частоты катастроф, то получится возрастающая зависимость частоты катастроф от времени. Этот эффект назвали «старением» микротрубочек — они как будто «портятся» со временем. Иначе говоря, «молодые» микротрубочки могут расти стабильно, а «старые» уже более склонны к разборке. Необычное распределение времен жизни микротрубочек хорошо аппроксимируется гамма-распределением, которое характеризует процессы с фиксированным количеством последовательных шагов. Поэтому возникла идея, что лучше всего результаты проведенного эксперимента описывает теория, согласно которой катастрофа микротрубочки происходит за три последовательных стадии, когда в ней накопились определенные дефекты неизвестной природы []. Эта гипотеза, исходно достаточно сомнительная, тем не менее существенно подогрела интерес к исследованию динамики микротрубочек на уровне отдельных димеров тубулина.
Рибосомы
По своей форме рибосомы напоминают сферу
В клетке рибосомы располагаются как свободно, так и в связи с эндоплазматической сетью. Чаще всего рибосомы бывают единичными, но возможны случаи, когда с молекулой информационной РНК ассоциируются две или более рибосом. Такую структуру называют полисомой. Полисомы состоят из одной молекулы иРНК и группы рибосом. Они выполняют функцию «считывания» информаци иРНК и создания полипептидных цепей в соответствии с нуклеотидной последовательностью.
Существуют два типа рибосом: прокариотические и эукариотические. Прокариотические характерны в основном для организмов-прокариотов, эукариотическое — для эукариотов. И те и другие имеют в своем составе все те же субъединицы и выполняют одни и те же функции. Примечательно, что рибосомы эукариот имеют больший размер, чем рибосомы прокариот.
Функция
Микротрубочки в клетке используются в качестве «рельсов» для транспортировки частиц. По их поверхности могут перемещаться мембранные пузырьки и митохондрии. Транспортировку по микротрубочкам осуществляют белки, называемые моторными. Это высокомолекулярные соединения, состоящие из двух тяжёлых (массой около 300 кДа) и нескольких лёгких цепей. В тяжёлых цепях выделяют головной и хвостовой домены. Два головных домена связываются с микротрубочками и являются собственно двигателями, а хвостовые — связываются с органеллами и другими внутриклеточными образованиями, подлежащими транспортировке.
Выделяют два вида моторных белков:
- цитоплазматические динеины;
- кинезины.
Динеины перемещают груз только от плюс-конца к минус-концу микротрубочки, то есть из периферийных областей клетки к центросоме. Кинезины, напротив, перемещаются к плюс-концу, то есть к клеточной периферии.
Перемещение осуществляется за счёт энергии АТФ. Головные домены моторных белков для этого содержат АТФ-связывающие участки.
Помимо транспортной функции, микротрубочки формируют центральную структуру ресничек и жгутиков — аксонему. Типичная аксонема содержит 9 пар объединённых микротрубочек по периферии и две полных микротрубочки в центре. Из микротрубочек состоят также центриоли и веретено деления, обеспечивающее расхождение хромосом к полюсам клетки при митозе и мейозе. Микротрубочки участвуют в поддержании формы клетки и расположения органоидов (в частности, аппарата Гольджи) в цитоплазме клеток.
Чего пока не может эксперимент и как помогает теория?
Обнаруженный феномен «старения» микротрубочек показал, что общепринятая, ставшая классической, модель ГТФ-«шапочки» — некоторое упрощение. Действительно, она только постулирует, что микротрубочка испытывает катастрофу, когда теряет свой стабилизирующий «колпачок», но не объясняет, как и почему это происходит, а также из-за чего же вообще микротрубочка может «стареть». Что за таинственные дефекты накапливаются внутри «стареющей» микротрубочки, приводя ее к катастрофе? Сколько их и в какой последовательности они должны проявляться? Может быть, речь идет о гидролизе отдельных молекул ГТФ внутри «шапочки» или о каком-то другом процессе, зависящем от не установленных пока событий совсем иной природы?
Естественно, исследователи хотели бы как можно тщательнее разглядеть «живые» микротрубочки, чтобы ответить на эти вопросы. Однако современный экспериментальный арсенал не позволяет это сделать. Мы можем или увидеть замороженную (обездвиженную) микротрубочку с нанометровым разрешением, например, с помощью электронного микроскопа, или проследить динамику микротрубочки со скоростью сотни кадров в секунду под оптическим микроскопом. К сожалению, невозможно получить соответствующие данные одновременно, чтобы четко их соотнести. Во многом по вине таких ограничений современной науке неизвестно, каков точный размер ГТФ-«шапочки» и как он меняется со временем, а также какую форму имеют концы микротрубочек и как она определяет их динамику.
На помощь экспериментам приходят теоретические методы исследования, в частности компьютерное моделирование. Оно может воссоздать микротрубочку с очень высоким пространственно-временным разрешением, правда, ценой неизбежных идеализаций и упрощений, адекватность которых нужно тщательно проверять (сравнивая результаты модельного и настоящего экспериментов). Идеальная компьютерная модель должна описывать все имеющиеся экспериментальные данные. Тогда на ее основе можно будет изучить механизмы наблюдаемого поведения микротрубочек и предсказать принцип действия белков, влияющих на динамику этих филаментов в клетках. Также станет возможным подбор химических соединений для управления поведением микротрубочек в медицинских целях.
На сегодняшний день создано множество моделей микротрубочек — от очень простых до весьма сложных. Самыми лучшими оказались наиболее детальные модели — молекулярные, которые учитывают, что микротрубочка состоит из многих протофиламентов и что ее структура дискретна (совокупность отдельных субъединиц — тубулинов). Первые такие модели стали появляться почти сразу после обнаружения динамической нестабильности в 1984 г. Работая с ансамблем взаимодействующих тубулинов, они воссоздают поведение микротрубочки как целого. Со времен первых молекулярных моделей накопилось много новых экспериментальных данных о микротрубочках. С тех пор уточнили их строение, измерили новые зависимости характеристик роста и укорочения от различных параметров, изучили поведение этих филаментов после разбавления тубулина, оценили размер ГТФ-«шапочки», открыли способность концов микротрубочек развивать тянущие и толкающие силы []. Это позволяло корректировать расчеты и все точнее задавать параметры взаимодействия тубулинов. Однако росли и требования к моделям, поскольку они должны непротиворечиво описывать весь набор имеющихся экспериментальных результатов. Таким образом, способы описания взаимодействия тубулинов совершенствовались и усложнялись. От простых моделей, где субъединицы либо взаимодействуют друг с другом, либо нет, перешли к так называемым молекулярно-механическим (самым современным и наиболее реалистичным). Они рассматривают молекулы тубулина как физические объекты, подчиняющиеся законам механики и движущиеся в поле тепловых соударений и потенциалов притяжения друг к другу []. В ранних молекулярно-механических расчетах динамики микротрубочек из-за ограниченной производительности компьютеров нельзя было подробно описать взаимодействие тубулинов на основе уравнений движения и с учетом тепловых колебаний. Однако эта цель оставалась очень притягательной для нашей команды, поскольку мы предполагали, что тепловые флуктуации играют существенную роль в динамике микротрубочек.
У животных
Центриоль
Основная статья: Центриоль
В большинстве клеток животных в течение интерфазы присутствует одна центросома, обычно располагающаяся возле ядра. В ней до S-периода содержится одна центриоль. В ходе S-периода достраивается вторая центриоль. Микротрубочки заякорены своими (-)-концами на центросоме, что предотвращает их полный распад. Обычно отходящие от центросомы микротрубочки характеризуются динамической нестабильностью: они медленно растут, а затем быстро распадаются почти целиком начиная с (+)-конца. Полярность микротрубочек важна для везикулярного транспорта: как правило, кинезины перемещают мембранные пузырьки с грузом к периферии клетки, двигаясь к (+)-концам, а динеины — в противоположном направлении.
Базальное тельце
В эпителиальных клетках животных, клетках сетчатки глаза и других нервных клетках — рецепторах, а также в клетках многих протистов ЦОМТы, содержащие центриоли, служат базальными тельцами ресничек и жгутиков.
У клеток зародышей животных и у большинства клеток самых разных тканей взрослого организма есть первичная ресничка, также обладающая базальным тельцем. Она играет важную роль в межклеточной сигнализации, миграции клеток и других процессах.
На базальных тельцах, как и на других ЦОМТах, происходит нуклеация микротрубочек, а заякоривание (-)-концов обеспечивает их стабилизацию. Базальные тельца обеспечивают рост ресничек и жгутиков, а также (вместе с корневыми структурами) закрепляют их на поверхности клетки.
Взаимодействие актин-миозиновой и тубулин-динеиновой комплекса
Актин-миозиновая и тубулин-динеиновая комплексы, как правило, в большинстве эукариотных клеток объединяются при функционировании в одну систему.
В частности, необходимые для запуска актин-миозиновой системы ионы кальция тормозят, а в высоких концентрациях и нарушают структурную организацию тубулин-динеиновой системы. Материал с сайта http://wiki-med.com
Постоянная смешанная микротрубочковая и актин-миозиновая система обнаружена в субмембранной области таких крайне специализированных образований, как кровяные пластинки млекопитающих, представляющие собой свободно циркулирующие в крови участки цитоплазмы полиплоидных клеток мегакариоцитов.
Помимо хорошо развитой в периферической гиалоплазме актин-миозиновой фибриллярной системы здесь имеется мощное кольцо микротрубочек, по-видимому, обеспечивающих поддержание формы этих структур.
Актин-миозиновая система кровяных пластинок играет важную роль в процессе свертывания крови.
Смешанные постоянные актин-миозиновая и тубулин-динеиновая системы, очевидно, широко распространены у высших простейших и, в частности, у инфузорий.
Однако в настоящее время они изучены преимущественно на уровне чисто морфо логического, ультраструктурного анализа. Функциональное взаимодействие названных двух основных механохимически: систем интенсивно исследуется у метазойных клеток в процессах митотического деления. Этот вопрос мы подробнее рассмотрим ниже, при описании процессов репродукции клеток.
Материал с сайта http://Wiki-Med.com
На этой странице материал по темам:
Динамическая нестабильность[ | код]
Микротрубочки являются динамическими структурами и в клетке постоянно полимеризуются и деполимеризуются. Центросома, локализованная вблизи ядра, выступает в клетках животных и многих протистов как центр организации микротрубочек (ЦОМТ): они растут от неё к периферии клетки. В то же время микротрубочки могут внезапно прекратить свой рост и укоротиться обратно по направлению к центросоме вплоть до полного разрушения, а затем вырасти снова. При присоединении к микротрубочке молекулы тубулина, несущие ГТФ, образуют «шапочку», которая обеспечивает рост микротрубочки. Если локальная концентрация тубулина падает, связанная с бета-тубулином ГТФ постепенно гидролизуется. Если полностью гидролизуется ГТФ «шапочки» на +-конце, это приводит к быстрому распаду микротрубочки. Таким образом, сборка и разборка микротрубочек связана с затратами энергии ГТФ.
Динамическая нестабильность микротрубочек играет важную физиологическую роль. Например, при делении клетки микротрубочки растут очень быстро и способствуют правильной ориентации хромосом и образованию митотического веретена.
Микрофиламенты
Каждый триплет центриоли связан со сферическими тельцами диаметром 75 нм – сателлитами; расходящиеся от них микротрубочки образуют центросферу.
Рис. 3-15. Клеточный центр (1) и структура центриоли (2). Клеточный центр образован парой центриолей (Ц), расположенных во взаимно-перпендикулярных плоскостях. Каждая Ц состоит из 9 связанных друг с другом триплетов (ТР) микротрубочек (МТ). С каждым ТР посредством ножек связаны сателлиты (С) – глобулярные белковые тельца, от которых отходят МТ.
В неделящейся клетке выявляется одна пара центриолей (диплосома), Которая обычно располагается вблизи ядра. Перед делением в S-периоде интерфазы происходит дупликация центриолей пары, причем под прямым углом к каждой зрелой (материнской) центриоли формируется новая (дочерняя), незрелая процентриоль, в которой вначале имеются лишь 9 единичных микротрубочек, позднее превращающихся в триплеты. Пары центриолей далее расходятся к полюсам клетки, а во время митоза они служат центрами образования микротрубочек ахроматинового веретена деления.
Рис. 3-16. Ресничка. 1 — продольный срез, 2 — поперечный срез. БТ — базальное тельце (образовано триадами микротрубочек), ЦОМТ — центр организации микротрубочек, БК — базальный корешок, ПЛ — плазмолемма, МТА — микротрубочка А, МТВ — микротрубочка В, ПМТ — периферические микротрубочки, ЦМТ — центральные микротрубочки, ЦО — центральная оболочка, ДР — динеиновые ручки, РС — радиаль-ные спицы, НМ — нексиновые мостики.
Дата добавления: 2014-01-07; Просмотров: 1381; Нарушение авторских прав?;
Микротрубочки — один из компонентов цитоскелета. Это цилиндрические структуры диаметром 22±2 нм и различной длины (от долей до сотен микрометров). Микротрубочки состоят в основном из одного белка — тубулина. Они хорошо видны в электронный микроскоп и могут быть выявлены, несмотря на свой малый диаметр, в световом микроскопе с помощью специального метода — иммунофлуоресценции.
Дело в том, что тубулины в клетках всех растений и животных сходны. Поэтому антитела (см. Антиген и антитело) к тубулину, полученному из мозга коров или свиней, будут «узнавать» микротрубочки в клетках практически любых организмов. Если к молекулам антител заранее химически присоединить маленькие молекулы светящихся при облучении сине-фиолетовыми лучами веществ — флуо-рохромы, то по распределению в клетках светящихся комплексов можно в микроскоп с флуоресцентной насадкой видеть расположение микротрубочек.
Микротрубочки образуют сеть в цитоплазме клеток, а во время деления формируют мито-тический аппарат (см. Митоз). Они входят в состав центриолей и базальных телец; жгутиков и ресничек. Цитоплазматические микротрубочки не стабильны, а находятся в динамическом равновесии с растворенным тубулином.
Расположение микротрубочек
Микротрубочки располагаются, как правило, в самых глубоких слоях примембранного цитозоля. Поэтому периферические микротрубочки надлежало бы рассматривать как часть динамичного, организующего микротрубочкового «скелета» клетки. Однако и сократимые, и скелетные фибриллярные структуры периферического цитозоля также связаны непосредственно с фибриллярными структурами основной гиалоплазмы клетки. В функциональном отношении периферическая опорно-сократимая фибриллярная система клетки находится в теснейшем взаимодействии с системой периферических микротрубочек. Это дает нам основание рассматривать последние как часть субмембранной системы клетки.