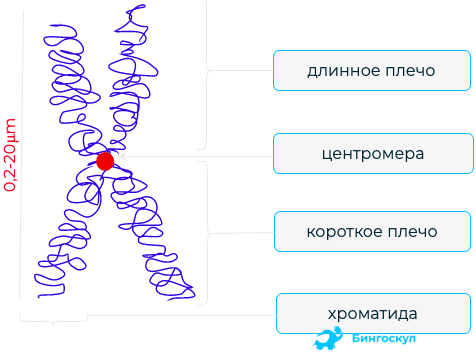
Компактизация днк
Содержание:
- История открытия хромосом
- История
- Необычные типы хромосом[ | код]
- Уровни организации молекулы ДНК
- Белки хроматина
- Бактериальные хромосомы
- История открытия хромосом
- Хромосомы человека
- Связь ДНК с белками ядра клетки
- Необычные типы хромосом
- Хромосомы человека
- Хромосомные аномалии
- Примечания
- Уровни компактизации хромосомной ДНК
- Гипотеза РНК мира
- Функция хромосом
- Суперскрученная молекула
- Репликация ДНК и ее строение
История открытия хромосом
Первые описания хромосом появились в статьях и книгах разных авторов в 70-х годах XIX века, и приоритет открытия хромосом отдают разным людям. Среди них такие имена, как И. Д. Чистяков (1873), А. Шнейдер (1873), Э. Страсбургер (1875), О. Бючли (1876) и другие. Чаще всего годом открытия хромосом называют 1882 год, а их первооткрывателем — немецкого анатома В. Флеминга, который в своей фундаментальной книге «Zellsubstanz, Kern und Zelltheilung» собрал и упорядочил сведения о них, дополнив результатами собственных исследований. Термин «хромосома» был предложен немецким гистологом Г. Вальдейером в 1888 году. «Хромосома» в буквальном переводе означает «окрашенное тело», поскольку оснóвные красители хорошо связываются хромосомами.
После переоткрытия в 1900 году законов Менделя потребовалось всего один-два года для того, чтобы стало ясно, что хромосомы при мейозе и оплодотворении ведут себя именно так, как это ожидалось от «частиц наследственности». В 1902 году Т. Бовери и в 1902—1903 годах У. Сеттон (Walter Sutton) независимо друг от друга выдвинули гипотезу о генетической роли хромосом.
Экспериментальное подтверждение этих идей было осуществлено в первой четверти XX века американскими учёными Т. Морганом, К. Бриджесом, А. Стёртевантом и Г. Мёллером. Объектом их генетических исследований послужила плодовая мушка D.melanogaster. На основе данных, полученных на дрозофиле, они сформулировали «хромосомную теорию наследственности», согласно которой передача наследственной информации связана с хромосомами, в которых линейно, в определённой последовательности, локализованы гены. Основные положения хромосомной теории наследственности были опубликованы в 1915 году в книге «The mechanism of mendelian heredity» (англ.).
В 1933 году за открытие роли хромосом в наследственности Т. Морган получил Нобелевскую премию по физиологии и медицине.
История
Первые наблюдения хромосом в ядре клетки, выполненные в 70-х годах 19 века И. Д. Чистяковым, О. Гертвигом, Страсбургером (E. Strasburger), положили начало цитологическому направлению в изучении хромосом. До начала 20 века это направление было единственным. Применение светового микроскопа позволило получить сведения о поведении хромосом в митотическом и мейотическом делениях (см. Мейоз, Митоз), факты о постоянстве числа хромосом у данного вида, специальных типах хромосом. В 20—40-х годах 20 века преимущественное развитие получило сравнительное морфологическое изучение хромосом у разных видов организмов, включая человека, с целью выяснения общих принципов их организации, особенностей индивидуальных хромосом и изменений их в процессе эволюции. В изучение этой проблемы особый вклад внесли отечественные ученые С. Г. Навашин, Г. А. Левитский, Л. Н. Делоне, П. И. Живаго, А. Г. Андрес, М. С. Навашин, А. А. П рокофъева-Бельговская, а также зарубежные — Хейтц (E. Heitz), Дарлингтон (С. D. Darlington) и др. С 50-х годов для исследования хромосом стал использоваться электронный микроскоп. Началось изучение морфологических изменений хромосом в процессе их генетического функционирования. В 1956 году Тио (H. J. Tjio) и Леван (A. Levan) окончательно установили число хромосом у человека, равное 46, описали их морфологические признаки в метафазе митоза. Значительный прогресс в изучении хромосом был достигнут в 70-х годах после разработки различных методов их окраски, позволивших выявить неоднородность структуры хромосом по длине в мета фазе деления клеток.
Сопоставление поведения хромосом в мейотическом делении с закономерностями наследования признаков (см. Менделя законы) положило начало цитогенетическим исследованиям. В конце 19 — начале 20 века Сеттоном (W. Sutton), Бовери (Th. Boveri), Уилсоном (Е. В. Wilson) были заложены основы хромосомной теории наследственности (см.), согласно которой гены локализованы в хромосомах и поведение последних при созревании гамет и их слиянии в момент оплодотворения объясняет законы передачи признаков в поколениях. Теория получила окончательное обоснование в цитогенетических экспериментах, проведенных на дрозофиле (см.) Т. Морганом и его учениками, которые доказали, что каждая хромосома есть группа генов, сцепленно наследуемых и расположенных в линейном порядке, что в мейозе осуществляется рекомбинация генов (см. Рекомбинация) гомологичных (идентичных) хромосом.
Изучение биохимической природы хромосом, начатое в 30—40-е годы 20 века, первоначально основывалось на цитохимическом качественном и количественном определении содержания ДНК, РНК и белков в ядре. С 50-х годов для этих целей стали применять фото- и спектрометрию (см. Спектрофотометрия), рентгеноструктурный анализ (см.) и другие физико-химические методы.
Необычные типы хромосом[ | код]
Микро- и макрохромосомы в метафазной пластинке курицы
B-хромосомы в метафазной пластинке сибирской косули Capreolus pygargus
Моноцентрическая (а) и голоцентрическая (b) хромосомы
Политенные хромосомы в клетке слюнной железы у представителя рода Chironimus из семейства комары-звонцы (Chironomidae)
Хромосома типа ламповых щеток из ядра ооцита тритона
Микрохромосомы | код
Основная статья: Микрохромосомы
У многих птиц и рептилий хромосомы в кариотипе образуют две чёткие группы: макрохромосомы и микрохромосомы. У некоторых видов микрохромосомы настолько мелкие и их так много, что невозможно отличить одну от другой. Микрохромосомы являются короткими по длине, но обогащёнными генами хромосомами. Например, кариотип курицы содержит 39 пар хромосом, 6 из которых относятся к макрохромосомам, а 33 — к минихромосомам. Макрохромосомы содержат две трети геномной ДНК, но только 25 % генов, в то время как микрохромосомы содержат оставшуюся треть геномной ДНК и 75 % генов. Таким образом, плотность генов в минихромосомах курицы в шесть раз выше, чем в макрохромосомах.
B-хромосомы | код
B-хромосомы — это добавочные хромосомы, которые имеются в кариотипе только у отдельных особей в популяции. Они часто встречаются у растений, описаны у грибов, насекомых и животных. Некоторые В-хромосомы содержат гены, часто это гены рРНК, однако не ясно, насколько эти гены функциональны. Наличие В-хромосом может влиять на биологические характеристики организмов, особенно у растений, где их наличие ассоциируется с пониженной жизнеспособностью. Предполагается, что В-хромосомы постепенно утрачиваются в соматических клетках в результате нерегулярности их наследования.
Голоцентрические хромосомы | код
Голоцентрические хромосомы не имеют первичной перетяжки, они имеют так называемый диффузный кинетохор, поэтому во время митоза микротрубочки веретена деления прикрепляются по всей длине хромосомы. Во время расхождения хроматид к полюсам деления у голоцентрических хромосом они идут к полюсам параллельно друг другу, в то время как у моноцентрической хромосомы кинетохор опережает остальные части хромосомы, что приводит к характерной V-образной форме расходящихся хроматид на стадии анафазы. При фрагментации хромосом, например, в результате воздействия ионизирующего излучения, фрагменты голоцентрических хромосом расходятся к полюсам упорядоченно, а не содержащие центромеры фрагменты моноцентрических хромосом распределяются между дочерними клетками случайным образом и могут быть утрачены.
Голоцентрические хромосомы встречаются у протист, растений и животных. Голоцентрическими хромосомами обладает нематода C. elegans.
Уровни организации молекулы ДНК
Различают четыре уровня, определяющих строение и морфологию этой гигантской молекулы:
- Первичный уровень, или структура – это порядок нуклеотидов в цепи.
- Вторичная структура представляет собой знаменитую «двойную спираль». Устоялось именно это словосочетание, хотя на самом деле подобная структура напоминает винт.
- Третичная структура образуется вследствие того, что между отдельными участками двухцепочечной закрученной нити ДНК возникают слабые водородные связи, придающие молекуле сложную пространственную конформацию.
- Четвертичная структура – это уже сложный комплекс ДНК с некоторыми белками и РНК. В такой конфигурации ДНК упакована в хромосомы в ядре клетки.

Белки хроматина
В структурной организации ДНП центральную роль играют специфические белки – гистоны.
Гистоны – это относительно небольшие по молекулярной массе белки, присутствуют в ядрах клеток эукариот. Гистоновые белки богаты остатками аминокислот аргинина и лизина, определяющими их щелочные свойства. Практически все гистоны одинаковы, среди них насчитывают 5–7 типов молекул, обладающих сходными свойствами. Гистоны – это структурные белки, выполняющие важную роль – упаковку ДНК. Например, в растянутом состоянии двойная спираль ДНК, содержавшаяся в хромосоме человека, имеет длину в среднем 4 – 5 см, а будучи спирализованной в хромосоме при участии гистонов, измеряется долями микрометра. По сравнению с остальными белками, присутствующими в клетке, количество гистонов в клетке очень велико – оно почти равно массе ДНК, содержащейся в ядре, что свидетельствует об их активном участии в структурировании хроматина. Известно также, что гистоновые белки являются регуляторами биосинтеза нуклеиновых кислот (и ДНК, и РНК). Гистоны синтезируются в цитоплазме, но затем транспортируются в ядро и там связываются с ДНК во время ее репликации. При этом синтез гистонов и ДНК синхронизирован. Молекулярный комплекс ДНК-гистоны имеет форму особых субъединиц – нуклеосом (от лат. nucleus – ядро и греч. soma – тело).
Кроме того, в состав хроматина входит значительное количество других белков, объединяемых общим названием «негистоновые белки».
Негистоновые белки в сравнении с гистонами, наоборот, очень разнообразны. В хроматине насчитывается нескольких сотен типов их молекул. Среди них – ферменты, обеспечивающие процессы репликации ДНК, транскрипции, а также некоторые белки ядерного матрикса и матрикса ядрышка. Полагают, что негистоновые белки хроматина выполняют и некоторые регуляторные функции. Именно негистоновые белки участвуют в формировании самых высоких уровней упаковки ДНК.
Хроматин – это самый существенный, основной компонент ядерного аппарата клетки; из него образованы хромосомы клеток эукариот.
Бактериальные хромосомы
Запрос «» перенаправляется сюда. На эту тему нужно создать отдельную статью (см. ).
Прокариоты (археи и бактерии, в том числе митохондрии и пластиды, постоянно обитающие в клетках большинства эукариот) не имеют хромосом в собственном смысле этого слова. У большинства из них в клетке имеется только одна макромолекула ДНК, замкнутая в кольцо (эта структура получила название нуклеоид). У ряда бактерий обнаружены линейные (не замкнутые в кольцо) макромолекулы ДНК. Помимо нуклеоида или линейных макромолекул, ДНК может присутствовать в цитоплазме прокариотных клеток в виде небольших замкнутых в кольцо молекул ДНК, так называемых плазмид, содержащих обычно незначительное, по сравнению с бактериальной хромосомой, число генов. Состав плазмид может быть непостоянен, бактерии могут обмениваться плазмидами в ходе парасексуального процесса.
Имеются данные о наличии у бактерий белков, связанных с ДНК нуклеоида, но гистонов у них не обнаружено.
История открытия хромосом
Первые описания хромосом появились в статьях и книгах разных авторов в 70-х годах XIX века, и приоритет открытия хромосом отдают разным людям. Среди них такие имена, как И. Д. Чистяков (1873), А. Шнейдер (1873), Э. Страсбургер (1875), О. Бючли (1876) и другие. Чаще всего годом открытия хромосом называют 1882 год, а их первооткрывателем — немецкого анатома В. Флеминга, который в своей фундаментальной книге «Zellsubstanz, Kern und Zelltheilung» собрал и упорядочил сведения о них, дополнив результатами собственных исследований. Термин «хромосома» был предложен немецким гистологом Г. Вальдейером в 1888 году. «Хромосома» в буквальном переводе означает «окрашенное тело», поскольку оснóвные красители хорошо связываются хромосомами.
После переоткрытия в 1900 году законов Менделя потребовалось всего один-два года для того, чтобы стало ясно, что хромосомы при мейозе и оплодотворении ведут себя именно так, как это ожидалось от «частиц наследственности». В 1902 году Т. Бовери и в 1902—1903 годах У. Сеттон (Walter Sutton) независимо друг от друга выдвинули гипотезу о генетической роли хромосом.
Экспериментальное подтверждение этих идей было осуществлено в первой четверти XX века американскими учёными Т. Морганом, К. Бриджесом, А. Стёртевантом и Г. Мёллером. Объектом их генетических исследований послужила плодовая мушка D.melanogaster. На основе данных, полученных на дрозофиле, они сформулировали «хромосомную теорию наследственности», согласно которой передача наследственной информации связана с хромосомами, в которых линейно, в определённой последовательности, локализованы гены. Основные положения хромосомной теории наследственности были опубликованы в 1915 году в книге «The mechanism of mendelian heredity» (англ.).
В 1933 году за открытие роли хромосом в наследственности Т. Морган получил Нобелевскую премию по физиологии и медицине.
Хромосомы человека
Нормальный кариотип человека представлен 46 хромосомами. Это 22 пары аутосом и одна пара половых хромосом (XY в мужском кариотипе и XX — в женском). В приведённой ниже таблице показано число генов и оснований в хромосомах человека.
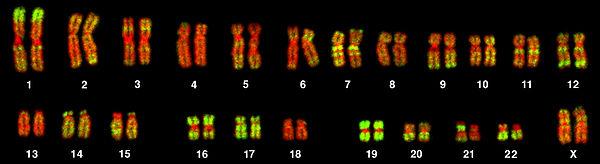
Изображение 46 (23 пар) хромосом женского кариотипа человека, полученное с помощью FISH с флуоресцентно-мечеными Alu-повторами. Alu-повторы показаны зелёным цветом, ДНК — красным. У человека самая длинная 1-я хромосома примерно в 5 раз длиннее самой короткой 21-й хромосомы.
| Хромосома | Всего оснований | Количество генов | Количество белок-кодирующих генов |
|---|---|---|---|
| 249250621 | 3511 | 2076 | |
| 243199373 | 2368 | 1329 | |
| 198022430 | 1926 | 1077 | |
| 191154276 | 1444 | 767 | |
| 180915260 | 1633 | 896 | |
| 171115067 | 2057 | 1051 | |
| 159138663 | 1882 | 979 | |
| 146364022 | 1315 | 702 | |
| 141213431 | 1534 | 823 | |
| 135534747 | 1391 | 774 | |
| 135006516 | 2168 | 1914 | |
| 133851895 | 1714 | 1068 | |
| 115169878 | 720 | 331 | |
| 107349540 | 1532 | 862 | |
| 102531392 | 1249 | 615 | |
| 90354753 | 1326 | 883 | |
| 81195210 | 1773 | 1209 | |
| 78077248 | 557 | 289 | |
| 59128983 | 2066 | 1492 | |
| 63025520 | 891 | 561 | |
| 48129895 | 450 | 246 | |
| 51304566 | 855 | 507 | |
| X-хромосома | 155270560 | 1672 | 837 |
| Y-хромосома | 59373566 | 429 | 76 |
Связь ДНК с белками ядра клетки
ДНК в ядрах клеток эукариот обычно находится в тесном взаимодействии с ядерными белками разных групп: основными (гистоновыми) и кислыми (негистоновыми). Ассоциируясь, ДНК и белки образуют единый нуклеопротеидный комплекс –дезоксирибонуклеопротеид (ДНП). Процентное соотношение сухого веса всех указанных компонентов ДНП таково: ДНК – 35–40 %, гистоновые белки – 30–50 %, негистоновые белки – 4–33 %, то есть 40 % сухого веса составляет ДНК и около 60 % – белки.
Термин «хроматин» (от греч. chroma – цвет, краска) был введен в 1880 году немецким гистологом Вальтером Флеммингом (1843–1905). Хроматин легко окрашивается ядерными красителями при исследовании клеточного препарата с помощью светового микроскопа.
Хроматином обычно называют дисперсное (деспирализированное) состояние хромосом в интерфазе клеточного ядра эукариот (в неделящейся клетке). Но с начала деления ядра молекулы ДНК уже спирализированы (упакованы) в хромосомы. Одна молекула ДНК (точнее – комплекс ДНП) представляет собой одну хромосому. Спирализацию, или упаковку, ДНК осуществляют преимущественно гистоны.
Необычные типы хромосом
Микро- и макрохромосомы в метафазной пластинке курицы
B-хромосомы в метафазной пластинке сибирской косули Capreolus pygargus
Моноцентрическая (а) и голоцентрическая (b) хромосомы
Политенные хромосомы в клетке слюнной железы у представителя рода Chironimus из семейства комары-звонцы (Chironomidae)
Хромосома типа ламповых щеток из ядра ооцита тритона
Микрохромосомы
Основная статья: Микрохромосомы
У многих птиц и рептилий хромосомы в кариотипе образуют две чёткие группы: макрохромосомы и микрохромосомы. У некоторых видов микрохромосомы настолько мелкие и их так много, что невозможно отличить одну от другой. Микрохромосомы являются короткими по длине, но обогащёнными генами хромосомами. Например, кариотип курицы содержит 39 пар хромосом, 6 из которых относятся к макрохромосомам, а 33 — к минихромосомам. Макрохромосомы содержат две трети геномной ДНК, но только 25 % генов, в то время как микрохромосомы содержат оставшуюся треть геномной ДНК и 75 % генов. Таким образом, плотность генов в минихромосомах курицы в шесть раз выше, чем в макрохромосомах.
B-хромосомы
B-хромосомы — это добавочные хромосомы, которые имеются в кариотипе только у отдельных особей в популяции. Они часто встречаются у растений, описаны у грибов, насекомых и животных. Некоторые В-хромосомы содержат гены, часто это гены рРНК, однако не ясно, насколько эти гены функциональны. Наличие В-хромосом может влиять на биологические характеристики организмов, особенно у растений, где их наличие ассоциируется с пониженной жизнеспособностью. Предполагается, что В-хромосомы постепенно утрачиваются в соматических клетках в результате нерегулярности их наследования.
Голоцентрические хромосомы
Голоцентрические хромосомы не имеют первичной перетяжки, они имеют так называемый диффузный кинетохор, поэтому во время митоза микротрубочки веретена деления прикрепляются по всей длине хромосомы. Во время расхождения хроматид к полюсам деления у голоцентрических хромосом они идут к полюсам параллельно друг другу, в то время как у моноцентрической хромосомы кинетохор опережает остальные части хромосомы, что приводит к характерной V-образной форме расходящихся хроматид на стадии анафазы. При фрагментации хромосом, например, в результате воздействия ионизирующего излучения, фрагменты голоцентрических хромосом расходятся к полюсам упорядоченно, а не содержащие центромеры фрагменты моноцентрических хромосом распределяются между дочерними клетками случайным образом и могут быть утрачены.
Голоцентрические хромосомы встречаются у протист, растений и животных. Голоцентрическими хромосомами обладает нематода C. elegans.
Хромосомы человека
Нормальный кариотип человека представлен 46 хромосомами. Это 22 пары аутосом и одна пара половых хромосом (XY в мужском кариотипе и XX — в женском). В приведённой ниже таблице показано число генов и оснований в хромосомах человека.
Изображение 46 (23 пар) хромосом женского кариотипа человека, полученное с помощью FISH с флуоресцентно-мечеными Alu-повторами. Alu-повторы показаны зелёным цветом, ДНК — красным. У человека самая длинная 1-я хромосома примерно в 5 раз длиннее самой короткой 21-й хромосомы.
| Хромосома | Всего оснований | Количество генов | Количество белок-кодирующих генов |
|---|---|---|---|
| 249250621 | 3511 | 2076 | |
| 243199373 | 2368 | 1329 | |
| 198022430 | 1926 | 1077 | |
| 191154276 | 1444 | 767 | |
| 180915260 | 1633 | 896 | |
| 171115067 | 2057 | 1051 | |
| 159138663 | 1882 | 979 | |
| 146364022 | 1315 | 702 | |
| 141213431 | 1534 | 823 | |
| 135534747 | 1391 | 774 | |
| 135006516 | 2168 | 1914 | |
| 133851895 | 1714 | 1068 | |
| 115169878 | 720 | 331 | |
| 107349540 | 1532 | 862 | |
| 102531392 | 1249 | 615 | |
| 90354753 | 1326 | 883 | |
| 81195210 | 1773 | 1209 | |
| 78077248 | 557 | 289 | |
| 59128983 | 2066 | 1492 | |
| 63025520 | 891 | 561 | |
| 48129895 | 450 | 246 | |
| 51304566 | 855 | 507 | |
| X-хромосома | 155270560 | 1672 | 837 |
| Y-хромосома | 59373566 | 429 | 76 |
Хромосомные аномалии
Анеуплоидия
Основная статья: Анеуплоидия
При анеуплоидии происходит изменение числа хромосом в кариотипе, при котором общее число хромосом не кратно гаплоидному хромосомному набору n. В случае утраты одной хромосомы из пары гомологичных хромосом мутантов называют моносомиками, в случае одной дополнительной хромосомы мутантов с тремя гомологичными хромосомами называют трисомиками, в случае утраты одной пары гомологов — нуллисомиками. Анеуплоидия по аутосомным хромосомам всегда вызывает значительные нарушения развития, являясь основной причиной спонтанных абортов у человека. Одной из самых известных анеуплоидий у человека является трисомия по хромосоме 21, которая приводит к развитию синдрома Дауна. Анеуплоидия характерна для опухолевых клеток, особенно для клеток сóлидных опухолей.
Полиплоидия
Основная статья: Полиплоидия
Изменение числа хромосом, кратное гаплоидному набору хромосом (n), называется полиплоидией. Полиплоидия широко и неравномерно распространена в природе. Известны полиплоидные эукариотические микроорганизмы — грибы и водоросли, часто встречаются полиплоиды среди цветковых, но не среди голосеменных растений. Полиплоидия клеток всего организма у многоклеточных животных редка, хотя у них часто встречается эндополиплоидия некоторых дифференцированных тканей, например, печени у млекопитающих, а также тканей кишечника, слюнных желёз, мальпигиевых сосудов ряда насекомых.
Хромосомные перестройки
Основная статья: Хромосомные перестройки
Хромосомные перестройки (хромосомные аберрации) — это мутации, нарушающие структуру хромосом. Они могут возникнуть в соматических и зародышевых клетках спонтанно или в результате внешних воздействий (ионизирующее излучение, химические мутагены, вирусная инфекция и др.). В результате хромосомной перестройки может быть утрачен или, наоборот, удвоен фрагмент хромосомы (делеция и дупликация, соответственно); участок хромосомы может быть перенесён на другую хромосому (транслокация) или он может изменить свою ориентацию в составе хромосомы на 180° (инверсия). Существуют и другие хромосомные перестройки.
Примечания
- ↑ Тарантул В. З. . Толковый биотехнологический словарь. — М.: Языки славянских культур, 2009. — 936 с. — 400 экз. — ISBN 978-5-9551-0342-6.
- Молекулярная биология клетки: в 3-х томах / Б. Альбертс, А. Джонсон, Д. Льюис и др. — М.-Ижевск: НИЦ «Регулярная и хаотическая динамика», Институт компьютерных исследований, 2013. — Т. I. — 808 с. — ISBN 978-5-4344-0112-8. — С. 309—336.
- , с. 13.
- , с. 9.
- ↑ , с. 12.
- (англ.). // Nobel Media AB 2013. Дата обращения 11 декабря 2013.
- , с. 29.
- ↑
- , с. 84—87.
- , с. 30.
- Рубцов Н. Б. . Методы работы с хромосомами млекопитающих: Учеб. пособие. — Новосибирск: Новосиб. гос. ун-т, 2006. — 152 с. — ISBN 5-94356-376-8.
- , с. 91.
- Разин С. В. . Хроматин: упакованный геном / С. В. Разин, А. А. Быстрицкий. — М.: БИНОМ: Лаборатория знаний, 2009. — 176 с. — ISBN 978-5-9963-0087-7.
- , с. 45—46.
- , с. 401—414.
- , с. 31.
- ↑
- (англ.). // Genome Reference Concortium. Дата обращения 18 апреля 2013.
- . // NCBI. Дата обращения 18 апреля 2013.
- Ensembl. (англ.). // The Ensembl project. Дата обращения 25 апреля 2013.
Уровни компактизации хромосомной ДНК

См. также: Сверхспирализация ДНК
Основу хромосомы составляет линейная макромолекула ДНК значительной длины. В молекулах ДНК хромосом человека насчитывается от 50 до 245 миллионов пар азотистых оснований. Суммарная длина ДНК из одной клетки человека составляет величину порядка двух метров. При этом типичное ядро клетки человека, которое можно увидеть только при помощи микроскопа, занимает объём около 110 мкм³, а митотическая хромосома человека в среднем не превышает 5—6 мкм. Подобная компактизация генетического материала возможна благодаря наличию у эукариот высокоорганизованной системы укладки молекул ДНК как в интерфазном ядре, так и в митотической хромосоме. Надо отметить, что у эукариот в пролиферирующих клетках осуществляется постоянное закономерное изменение степени компактизации хромосом. Перед митозом хромосомная ДНК компактизуется в 105 раз по сравнению с линейной длиной ДНК, что необходимо для успешной сегрегации хромосом в дочерние клетки, в то время как в интерфазном ядре для успешного протекания процессов транскрипции и репликации хромосоме необходимо декомпактизоваться. При этом ДНК в ядре никогда не бывает полностью вытянутой и всегда в той или иной степени упакована. Так, расчётное уменьшение размера между хромосомой в интерфазе и хромосомой в митозе составляет всего примерно 2 раза у дрожжей и 4—50 раз у человека.
Упаковка ДНК в хроматин обеспечивает многократное сокращение линейных размеров ДНК, необходимое для размещения её в ядре. Она происходит в несколько этапов. Наиболее изученными являются три первых уровня упаковки: (1) накручивание ДНК на нуклеосомы с образованием нуклеосомной нити диаметром 10 нм, (2) компактизация нуклеосомной нити с образованием так называемой 30-нм фибриллы и (3) сворачивание последней в гигантские (50 — 200 тысяч п. н.) петли, закреплённые на белковой скелетной структуре ядра — ядерном матриксе.
Одним из самых последних уровней упаковки в митотическую хромосому некоторые исследователи считают уровень так называемой хромонемы, толщина которой составляет около 0,1—0,3 мкм. В результате дальнейшей компактизации диаметр хроматиды достигает ко времени метафазы 700 нм. Значительная толщина хромосомы (диаметр 1400 нм) на стадии метафазы позволяет, наконец, увидеть её в световой микроскоп. Конденсированная хромосома имеет вид буквы X (часто с неравными плечами), поскольку две хроматиды, возникшие в результате репликации, соединены между собой в районе центромеры (подробнее о судьбе хромосом при клеточном делении см. статьи митоз и мейоз).
Гипотеза РНК мира
Концепция РНК мира заключается в том, что когда-то очень давно молекула РНК могла выполнять функцию как молекулы ДНК, так и белков.
В живых организмах практически все процессы происходят благодаря ферментам белковой природы. Белки, однако, не могут самореплицироваться и синтезируются в клетки на основании информации, заложенной в ДНК. Но и удвоение ДНК происходит только благодаря участию белков и РНК. Следовательно, образуется замкнутый круг, из-за которого в рамках теории возникновения жизни спонтанное возникновение такой сложной системы маловероятно.
В начале 1980-х годов в лаборатории ученых Чека и Олтмена (обладатели нобелевской премии по химии) в США была открыта каталитическая способность РНК. РНК-катализаторы были названырибозимами (см. Рис. 11).
Рис. 11. Структура рибозимомолекулы РНК, выполняющей функцию катализа
Оказалось, что активный центр рибосом тоже содержит большое количество рибосомных РНК. Также РНК способны создавать двойную цепочку и самореплицироваться. То есть РНК могли существовать полностью автономно, катализируя метаболические реакции, например синтеза новых рибонуклеатидов, и самовоспроизводясь, сохраняя из поколения в поколение каталитические свойства. Накопление случайных мутаций привело к появлению РНК, катализирующих синтез определенных белков, являющихся более эффективными катализаторами, в связи с чем эти мутации закреплялись в ходе естественного отбора. Также возникли специализированные хранилища генетической информации – молекула ДНК, а РНК стала посредником между ДНК и белками.
Функция хромосом
Функция хромосом заключается:
1) в хранении наследственной информации. Хромосомы являются носителями генетической информации;
2) передаче наследственной информации. Наследственная информация передается путем репликации молекулы ДНК;
3) реализации наследственной информации. Благодаря воспроизводству того или иного типа и-РНК и, соответственно, того или иного типа бел- ка осуществляется контроль над всеми процессами жизнедеятельности клетки и всего организма.
Таким образом, хромосомы с заключенными в них генами обусловливают непрерывный ряд воспроизведения.
Генетические функции хромосом до начала 40-х годов XX века большинство исследователей связывали с белками. Как писал русский биолог, генетик Николай Константинович Кольцов, «трудно было признать, что за такой простенькой молекулой, как ДНК, столь сложные функции». Однако в дальнейшем именно ДНК была идентифицирована как генетический материал всех растений, животных, микроорганизмов и большинства вирусов.
Хромосомы осуществляют сложную координацию и регуляцию процессов в клетке вследствие заключенной в них генетической информации, обеспечивающей синтез первичной структуры белков-ферментов.
У каждого вида в клетках находится определенное количество хромосом. Они являются носителями генов, определяющих наследственные свойства клеток и организмов вида. Ген – это участок молекулы ДНК хромосомы, на котором синтезируются различные молекулы РНК (трансляторы генетической информации). Ген является функционально неделимой единицей генетического материала и представлен в виде определенного числа (не менее трех) линейно расположенных нуклеотидов. Именно последовательность нуклеотидов в цепях ДНК определяет код генетической информации, способной транскрибироваться (списываться, как с матрицы) на молекулы иРНК, а они, в свою очередь, транслируют (передают) в соответствии с кодом последовательность включения аминокислот в синтезирующуюся полимерную молекулу белка по принципу «один ген – один белок». Количество нуклеотидов в хромосомах огромно. Например, гены, входящие в состав хромосом человека, могут содержать до двух миллионов пар нуклеотидов.
Суперскрученная молекула
Сверхспирализованная форма ДНК возникает, когда двухцепочечная молекула совершает дополнительное закручивание. Оно может быть направлено по часовой стрелке (положительно) либо против (в этом случае говорят об отрицательной сверхспирализации). ДНК большинства организмов суперскручена отрицательно, то есть против основных витков «двойной спирали».
В результате образования дополнительных петель – супервитков – ДНК приобретает сложную пространственную конфигурацию. В клетках эукариот этот процесс происходит с формированием комплексов, в которых ДНК отрицательно навивается на гистоновые белковые комплексы и принимает вид нити с бусинами-нуклеосомами. Свободные участки нити называются линкерами. В поддержании суперскрученной формы молекулы ДНК принимают участие и негистоновые белки, а также неорганические соединения. Так образуется хроматин – вещество хромосом.
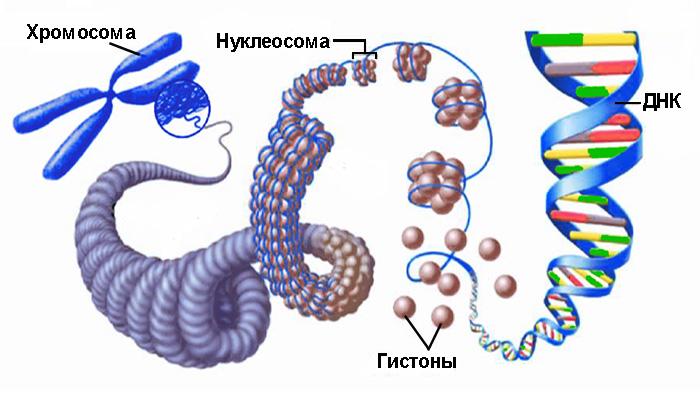
Хроматиновые нити с нуклеосомными бусинами способны к дальнейшему усложнению морфологии в процессе, называемом конденсацией хроматина.
Репликация ДНК и ее строение
И первичная, и вторичная структуры ДНК возникают в ходе явления, называемого репликацией – образования из материнской макромолекулы двух идентичных ей «двойных спиралей». При репликации исходная молекула расплетается, и на освободившихся одиночных цепочках происходит наращивание комплементарных оснований. Поскольку половинки ДНК антипараллельны, этот процесс протекает на них в разных направлениях: по отношению к материнским цепочкам от 3’-конца к 5’-концу, то есть новые цепочки растут в направлении 5’ → 3’. Лидирующая цепь синтезируется непрерывно в сторону репликационной вилки; на отстающей цепи синтез совершается от вилки отдельными участками (фрагменты Оказаки), которые затем сшивает между собой особый фермент – ДНК-лигаза.

Пока продолжается синтез, уже сформированные концы дочерних молекул претерпевают винтообразное закручивание. Затем, еще до окончания репликации новорожденные молекулы начинают образовывать третичную структуру в процессе, именуемом сверхспирализацией.