Спирали днк: основные понятия, структура, выполняемые функции и генетика
Содержание:
- Создание модели
- Химия и генетика
- Пытливый исследователь
- Древние перспективы
- Лидирующая научная гипотеза о том, как появилась жизнь на Земле
- Как была открыта структура ДНК
- ДНК не по канону
- Эластичность
- Разница между ДНК и РНК-полимеразой
- «Мир РНК»
- Рыбалка на благо науки
- Как хорошо уметь считать!
- Семейства ДНК-полимераз
- Уровни структурной организации
- Надежда на быстрый успех
- Структура двойной спирали ДНК
- Немного теории для не биологов
- Post Scriptum
Создание модели
В конце 1970-х годов в качестве потенциального решения проблем репликации ДНК в плазмидах и хроматине были кратко рассмотрены альтернативные неспиральные модели. Однако они были отвергнуты в пользу двойной модели, изображающей виток спирали ДНК, из-за последующих экспериментальных достижений, таких как рентгеновская кристаллография ДНК-дуплексов. Кроме того, не двойные спиральные модели в настоящее время не принимаются основным научным сообществом.
Одноцепочечные нуклеиновые кислоты (ssDNA) не принимают спиральную форму и описываются такими моделями как случайная катушка или червоподобная цепь.
ДНК является относительно жестким полимером, типично смоделированным как червоподобная цепь. Модельная жесткость важна для циркуляризации ДНК и ориентации связанных с ней белков относительно друг друга, а гистерезисная осевая жесткость важна для обертывания ДНК и циркулирования и взаимодействия белков
Сжатие-удлинение относительно неважно при отсутствии высокого напряжения
Химия и генетика
ДНК в растворе не принимает жесткую структуру, но постоянно меняет конформацию из-за тепловой вибрации и столкновения с молекулами воды, что делает невозможным применение классических мер жесткости. Следовательно, изгибная жесткость ДНК измеряется длиной персистентности, определяемой как «длина ДНК, по которой усредненная по времени ориентация полимера становится некоррелированной по коэффициенту».
Это значение может быть точно измерено с помощью атомного силового микроскопа для непосредственного изображения молекул ДНК различной длины. В водном растворе средняя постоянная длина составляет 46-50 нм или 140-150 пар оснований (диаметр ДНК 2 нм), хотя она может значительно различаться. Это делает ДНК умеренно жесткой молекулой.
Длительность продолжения участка ДНК сильно зависит от ее последовательности, и это может привести к значительным изменениям. Последние в большинстве своем обусловлены энергией штабелирования и фрагментами, которые распространяются на минорные и крупные канавки.
Пытливый исследователь
Ученый менял способы промывки лейкоцитов, реагенты и способы очистки. Так выяснилось, что это вещество — не белок и не жир. Биохимия только зарождалась, химический анализ был делом непростым, долгим и очень трудоемким. Но Фридрих Мишер провел его и выяснил, что составляющие ДНК — это углерод, кислород и азот. Но что более странно – в веществе присутствовал в больших концентрациях фосфор. На тот момент химия не знала соединений подобного состава, и Мишер понял, что он открыл что-то особенное. Гоппе-Зейлер (наставник) поддержал его, и в 1871 году труды были опубликованы и сопровождались рецензией уважаемого химика того времени. Дальнейшее изучение показало кислотные свойства вещества, и именно тогда оно стало называться нуклеиновой кислотой. Хотя, по правде, первооткрывателю это не нравилось, и в своих работах он продолжал называть ДНК нуклеином.

Древние перспективы
Идеальные случаи, связанные с превосходной сохранностью археологического материала, приводят к тому, что условная фаланга пальца или бедренная кость, найденные где-то в Сибири, становятся прорывом в понимании истории человечества, заставляющим пересмотреть всю его историю или даже опровергнуть казавшуюся незыблемой теорию его происхождения.
Такие находки и такие исследования приводят к появлению не только великолепных научных публикаций, но и позволяют зародить надежду на возрождение прошлого. Увы, современные естественнонаучные методы позволяют лишь приподнять занавесь древней истории нашей планеты, но воскресить его вряд ли смогут.
Лидирующая научная гипотеза о том, как появилась жизнь на Земле
Итак, лидирующая гипотеза в возникновении жизни на Земле — больше не «мир РНК».
Найдено связующее звено — тиоуридин — которое способно привести к возникновению как ДНК, так и РНК одновременно, создав строительные блоки для первых клеток с генами.
А именно первые микробы — одноклеточные живые организмы — и стали самой первой формой жизни на Земле. Это случилось примерно 4 миллиарда лет назад (исследователи обнаружили следы первых микробов в скалах, которым 3 миллиарда 700 тысяч лет).
И после миллиардов лет эволюции в результате мутаций появился новый вид — homo sapiens, человек разумный. Это мы с вами.
Это событие произошло меньше полумиллиона лет назад, хотя приматы (к которым относится человек) отделились как ветвь от других хордовых примерно 85 миллионов назад (85 000 000 лет). Основываясь исследованиях археологии и генома человека, ученые датировали появление вида homo sapiens примерно 200 тысяч лет назад (200 000).
Как на Земле появился человек
То есть, человечество — относительно очень молодой вид. Его возраст — 1/425 возраста приматов как отдельного семейства хордовых (позвоночных).
Не путайте понятия «возникновение жизни на Земле» (первые живые организмы — микробы) и «возникновение разумной жизни» (единственный известный нам биологический вид, доживший до наших дней — это мы, homo sapiens, человек разумный).
Жизнь на Земле появилась очень давно, а человек — совсем недавно. Произошло это в результате процесса эволюции.
Жизнь на Земле останется и тогда, когда исчезнет человек как вид. А это вполне возможно, как мы выяснили на примере вымерших динозавров. Их тоже погубило изменение климата.
И именно то, что с планеты исчезли динозавры, дало толчок развитию хордовых, в том числе и приматов, примерно 66 миллионов лет назад.
Увлекательно? Читайте подробности по ссылкам ниже.
 Прочтите также:
Прочтите также:
- Как появилась жизнь на Земле. Секретные материалы ученых
- Что случилось 66 миллионов лет назад, когда вымерли динозавры
- Двигатель эволюции — не борьба за выживание, а взаимопомощь, считают ученые
- Как на Земле появился человек? Событие, которое изменило жизнь на Земле
Как была открыта структура ДНК
Над открытием структуры ДНК в середине ХХ века бились многие ученые. Но только трое из них: Джеймс Уотсон, Фрэнсис Крик и Морис Уилкинс в 1962 году были удостоены Нобелевской премии по физиологии и медицине.
Начало истории
Джеймс Уотсон родился в 1928 году. В момент открытия структуры ДНК ему было всего 25 лет. В 1947-1951 годах Джеймс Уотсон учился в магистратуре и аспирантуре Индианского университета. Под руководством итальянского ученого-рентгенолога Сальвадора Лурии он написал диссертацию о воздействии рентгеновских лучей на размножение бактериофагов. В 1950 году Джеймс Уотсон получил докторскую степень. Изучая строение бактериофагов, Джеймс Уотсон, используя генетические методы, пытался определить структуру ДНК. Однако после доклада физика из Лондонского королевского колледжа Мориса Уилкинса, он понял, что для открытия структуры ДНК нужно использовать метод рентгеноструктурного анализа.
С целью изучения этого метода Джеймс Уотсон в 1951 году поступил в Кавендишскую лабораторию Кембриджского университета, где начал изучать структуру белков. Там он познакомился с физиком Фрэнсисом Криком, который интересовался биологией и был силен в теории кристаллографии.
Основные события
Параллельно работам в Кавендишской лаборатории Кембриджского университета над расшифровкой структуры ДНК работали сотрудники Лондонского королевского колледжа Морис Уилкинс и Розалинда Франклин. Розалинда Франклин ( рис.1) прекрасно владела методом рентгеноструктурного анализа и ее рентгенограммы ДНК отличались высоким качеством и четкостью.

Рис.1. Розалинда Франклин
Пытался разгадать структуру ДНК и нобелевский лауреат химик Лайнус Полинг. Однако его модели предполагали, что ДНК состоит из трех цепей.
В декабре 1952 года Морис Уилкинс без согласия Розалинды Франклин продемонстрировал Джеймсу Уотсону рентгенограмму №51, которая отличалась высокой четкостью. По одним данным ее выполнила Розалинда Франклин, по другим – Раймонд Гослинг. Эта рентгенограмма помогла Джеймсу Уотсону и Фрэнсису Крику построить модель структуры ДНК (рис.2).
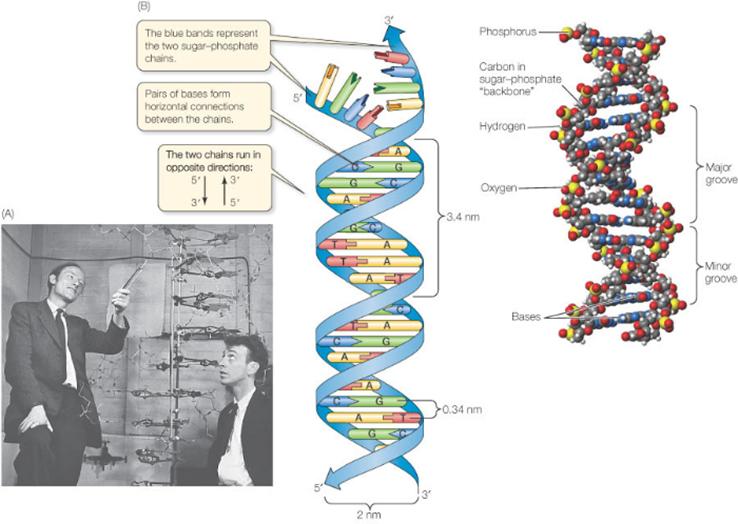
Рис.1. Фрэнсис Крик (слева) и Джеймс Уотсон возле модели ДНК
«Мой рот открылся, и мой пульс начал биться» – написал Уотсон в своей знаменитой книге «Двойная спираль». Это была единственная информация, которая была нужна ему и Френсису Крику, чтобы составить точную модель структуры ДНК. Рентгенограмма 51 была доказательством того, что спиральная структура ДНК имела две нити, прикрепленные в середине фосфатными основаниями. В апрельском номере журнала Nature за 1953 год они опубликовали краткую статью, в которой описывали структуру ДНК и даже поместили набросок ее структуры. В том же номере журнала также была опубликована статья Мориса Уилкинса и Розалинды Франклин. Но, как отмечают последующие исследователи этой почти детективной истории, статья Джеймса Уотсона и Фрэнсиса Крика была краткой и четкой. Статья Розалинды Франклин и Мориса Уилкинса была менее строгой и понятной.
Конец истории
В 1958 году Розалинда Франклин умерла. У нее был диагносцирован рак. Возможно, сказалось длительное рентгеновское облучение при работе над рентгенограммами ДНК, возможно – огорчение, что другие воспользовались ее трудами без ее ведома.
В 1962 году за открытие структуры ДНК Джеймс Уотсон, Фрэнсис Крик и Морис Уилкинс были удостоены Нобелевской премии по физиологии и медицине.
ДНК не по канону
Мы привыкли думать о ДНК как о двойной спирали, в которой азотистые основания нуклеотидов на противоположных цепях однозначно соответствуют друг другу: аденин — тимину, гуанин — цитозину. Эта, безусловно, фундаментальная особенность нуклеиновых кислот лежит в основе механизмов наследственности. Именно благодаря ей становятся возможными удвоение и корректирование ДНК, а также реализация генетической информации в структуре РНК и белков.
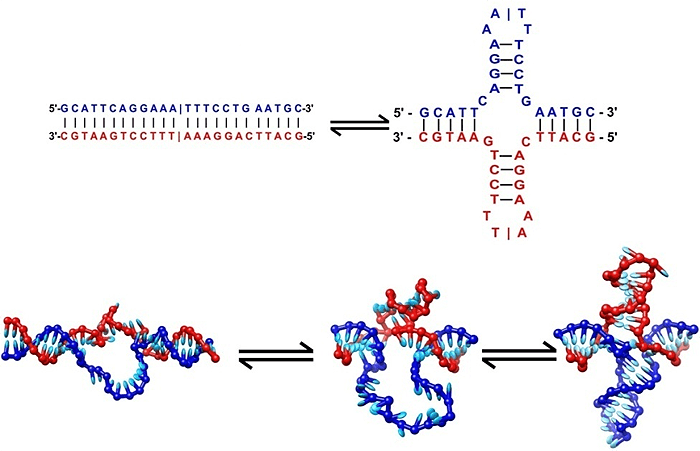
Рисунок 1. Формирование крестоообразных структур ДНК.

Рисунок 2. Двойная (а), тройная (б) и четверная (в) спирали ДНК.
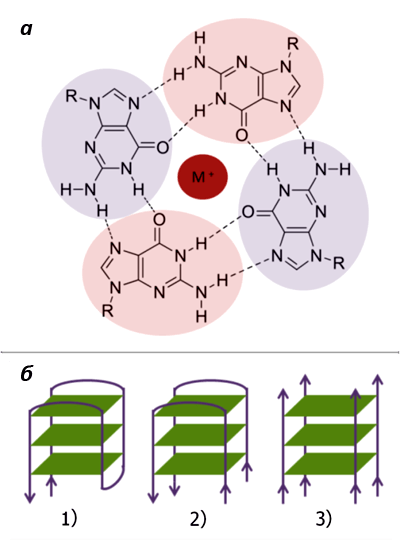
Рисунок 3. «G-ДНК». а. Строение G-тетрады; М+ — одновалентный катион. б. Формирование G-квадруплекса из 1) одной, 2) двух и 3) четырех нитей ДНК.
Способность гуанина к самоассоциации была обнаружена еще в конце 19 века. И только в 1962 году удалось установить, что в растворах он образует агрегаты из четырех молекул (называемых G-квартетом, или G-тетрадой) . Такие тетрады скрепляются между собой неканоническими (то есть, не предусмотренными в модели Уотсона — Крика) водородными связями, называемыми «хугстиновскими» — по фамилии их первооткрывателя . При этом входящие в них гуанины располагаются в одной плоскости и нуждаются в стабилизации моновалентными катионами (например, K+ или Na+) (рис. 3а).
Содержащие гуанин нуклеиновые кислоты в растворе могут образовывать такие структуры из одной, двух или четырех различных нитей (рис. 3б). Однако стабильными они будут лишь в том случае, когда три и более G-тетрады сгруппируются в плотную «стопку», «подперев» друг друга межплоскостными стекинг-взаимодействиями и сформировав тем самым G-квадруплекс (G4-структуру). А для этого, в свою очередь, необходима «счастливая встреча» четырех полигуаниновых участков (GGGn), находящихся либо на одной, либо на разных молекулах ДНК или РНК .
Нанопроводники
Интересно, что полигуаниновые цепи ДНК в растворе образуют четырехспиральные жгуты, для которых, к слову, зафиксирована способность эффективно проводить ток. Это свойство создает перспективу их использования в качестве нанопроводников .
Эластичность
Более длинные участки ДНК энтропически эластичны при растяжении. Когда ДНК находится в растворе, она подвергается непрерывным структурным изменениям из-за энергии, доступной в термальной ванне растворителя. Это связано с тепловыми колебаниями молекулы ДНК в сочетании с постоянными столкновениями с молекулами воды. По энтропийным причинам более компактные расслабленные состояния являются термически более доступными, чем растянутые состояния, и поэтому молекулы ДНК почти повсеместно встречаются в запутанных «расслабленных» молекулярных моделях. По этой причине одна молекула ДНК будет растягиваться под действием силы, выправляя ее. Используя оптические пинцеты, энтропийное растягивающее поведение ДНК изучалось и анализировалось с точки зрения физики полимеров, и было обнаружено, что ДНК ведет себя в основном как червеподобная цепная модель Кратки-Порода в физиологически доступных энергетических масштабах.
При достаточном натяжении и положительном крутящем моменте ДНК, как считается, подвергается фазовому переходу, при этом основы расходятся наружу, а фосфаты перемещаются в середину. Эта предложенная структура для перенапряженной ДНК была названа ДНК P-формы в честь Линуса Полинга, который изначально представлял ее как возможную структуру ДНК.
Доказательства механического растяжения ДНК в отсутствие наложенного крутящего момента указывают на переход или переходы, ведущие к дальнейшим структурам, которые обычно называются S-формами. Эти структуры еще не были окончательно охарактеризованы из-за сложности выполнения изображений с разрешением атомного резонатора в растворе при приложении силы, хотя многие компьютерные симуляционные исследования были сделаны. Предлагаемые структуры S-ДНК включают в себя те, которые сохраняют укладку базовой пары и водородную связь (обогащенную GC).

Разница между ДНК и РНК-полимеразой
Определение
ДНК-полимераза: ДНК-полимераза — это фермент, который синтезирует новые молекулы ДНК из нуклеотидов ДНК в процессе, называемом репликация ДНК.
РНК-полимераза: РНК-полимераза — это фермент, ответственный за синтез молекул РНК из ДНК в процессе, называемом транскрипцией.
использование
ДНК-полимераза: ДНК-полимераза используется в репликации ДНК.
РНК-полимераза: РНК-полимераза используется в транскрипции.
Товар
ДНК-полимераза: ДНК-полимераза синтезирует двухцепочечную молекулу ДНК.
РНК-полимераза: РНК-полимераза синтезирует одноцепочечную молекулу РНК.
Нуклеотиды
ДНК-полимераза: ДНК-полимераза использует нуклеотиды ДНК для синтеза новой цепи.
РНК-полимераза: РНК-полимераза использует РНК-нуклеотиды для синтеза новой цепи.
Активность в клеточном цикле
ДНК-полимераза: ДНК-полимераза активна во время S-фазы интерфазы.
РНК-полимераза: РНК-полимераза активна в течение G1 и G2 фазы фазы роста.
Ферментное связывание
ДНК-полимераза: Связывание ДНК-полимеразы с цепью требует репликационной вилки, которая производится ДНК-гиразой.
РНК-полимераза: Связывание РНК-полимеразы с цепью требует, чтобы ее голофермент распознал промотор гена.
грунтовка
ДНК-полимераза: ДНК-полимераза требует праймера для инициации репликации.
РНК-полимераза: РНК-полимераза не требует праймера для инициации транскрипции.
Экзонуклеазная активность
ДНК-полимераза: ДНК-полимераза обладает экзонуклеазной активностью.
РНК-полимераза: РНК-полимераза лишена экзонуклеазной активности.
относительное удлинение
ДНК-полимераза: ДНК-полимераза синтезирует всю хромосому.
РНК-полимераза: РНК-полимераза останавливает синтез в месте терминации транскрипции.
скорость
ДНК-полимераза: Кишечная палочка ДНК-полимераза добавляет 1000 нуклеотидов в секунду.
РНК-полимераза: Кишечная палочка РНК-полимераза добавляет 40-80 нуклеотидов в секунду.
Частота ошибок
ДНК-полимераза: ДНК-полимераза имеет меньшую частоту ошибок.
РНК-полимераза: РНК-полимераза имеет высокую частоту ошибок по сравнению с ДНК-полимеразой.
Типы
ДНК-полимераза: Pol I и Pol III — это два типа ДНК-полимераз, которые отвечают за 80% репликации ДНК в клетке.
РНК-полимераза: Эукариоты содержат РНК-полимеразы I-V. Прокариоты содержат только один тип РНК-полимераз.
Заключение
ДНК и РНК-полимераза являются двумя ферментами, ответственными за синтез молекул ДНК и РНК из генетического материала внутри ядра. ДНК-полимераза используется в репликации ДНК, а РНК-полимераза участвует в транскрипции. Основное различие между ДНК и РНК-полимеразой заключается в их функции и требованиях.
«Мир РНК»
Нуклеиновые кислоты открыл в 1868 году швейцарский ученый Иоганн Фридрих Мишер. Термин «нуклеин» был употреблен, поскольку эти вещества первоначально обнаружились в ядре клетки (ядро по-латыни nucleus). Но позднее оказалось, что клетки бактерий, не имеющие ядра, содержат те же кислоты.
Рибонуклеиновая кислота (РНК) — это одна из трех основных макромолекул, наряду с ДНК и белками, содержащихся в клетках живых организмов. К образованию клеточных РНК приводит процесс, называемый транскрипцией, — синтез РНК на матрице ДНК, осуществляемый посредством специальных ферментов, РНК-полимеразы. Матричные РНК (мРНК), в свою очередь, участвуют в процессе трансляции — синтезе белка на матрице мРНК с помощью рибосом. Остальные разновидности РНК после транскрипции модифицируются и выполняют различные функции, зависящие от их типа, скажем, доставки аминокислот к месту синтеза белка.
Молекулы РНК входят в состав некоторых ферментов (таких, как теломераза), но отдельные виды обладают собственной активностью. Так, они могут вносить разрывы в другие молекулы РНК или, напротив, «склеивать» между собой два РНК-фрагмента.
Из РНК состоят геномы некоторых вирусов. Поскольку эти молекулы способны одновременно выполнять функции и носителя информации, и катализатора химических реакций, была выдвинута гипотеза, согласно которой они стали первыми на Земле сложными полимерами.
Так называемая «гипотеза РНК-мира» гласит, что в начале эволюции РНК стала катализатором для синтеза других аналогичных молекул, а затем и ДНК. Но постепенно в большинстве структур РНК заменил белок.
Рыбалка на благо науки
Возвратившись в Базель, и заняв пост заведующего кафедры физиологии университета, в котором учился, Мишер продолжил научные изыскания. Объектом для изучения ДНК он выбрал молоки лосося. Кстати, и сегодня молоки лососевых используются для получения ДНК в больших количествах. В Рейне в те времена лосось водился в избытке, и Фридрих совмещал приятное времяпровождение с удочкой с работой по добыче материала для исследований. В своих работах о молоках (1874 г.) он и не предполагал роли ДНК в наследовании признаков, а связывал это вещество с процессами оплодотворения. От изучения молок лососевых он перешел к изучению их физиологии.
Позже ученый приступил к исследованию вопросов здорового рациона для заключенных тюрем. Он даже написал поваренную книгу. Затем он основал институт анатомии и физиологии (Базель), внес вклад в изучение роли кровяных клеток в дыхании. А про его нуклеотид надолго забыли.

Как хорошо уметь считать!
После запуска этих машин и получения первичных, «сырых» данных палеогенетики молитвенно складывают руки, ожидая результатов, которые будут получены группой людей, объединенных специальностью биоинформатика . Именно биоинформатики являются последним звеном при работе с древней ДНК, извлеченной из археологического или палеонтологического материала.
Между нами, эта их особенность дает им уверенность в своем превосходстве над остальными звеньями научного эксперимента, что часто ведет к торможению многих блестящих научных проектов. Однако с этим приходится мириться, поскольку современная популяционная генетика невозможна без использования биоинформатических методов, и на Западе это десятки людей, связанных одной целью. Обработка данных секвенирования древней ДНК — не самое тривиальное занятие: несмотря на значительные усилия, в данных по-прежнему остается большой процент современной ДНК, а также неидентифицируемого генетического мусора, который нуждается в тщательной очистке, с привлечением математического аппарата. Удивительно, но именно постмортальные модификации нуклеотидных последовательностей древней ДНК и небольшая длина выделенных из биологического материала ДНК-фрагментов служат превосходным маркером при работе с древней ДНК, позволяя отфильтровать весь современный генетический мусор (рис. 3).

Рисунок 3. Тест на контаминацию современным генетическим материалом .
Фактически фильтрация данных древней ДНК — наиболее важная часть биоинформатической работы: именно на этой стадии отсекаются все те последовательности, которые впоследствии могут привести к научной ошибке. Далее следуют стандартные этапы по определению митохондриальных гаплогрупп, Y-хромосомных гаплотипов, поиску древних геномных мутаций и даже сборке нового генома (например, используя уже известные геномы современных людей в качестве «каркаса»).
Семейства ДНК-полимераз
На основании своей структуры ДНК-полимеразы могут быть разбиты на семь различных семейств: A, B, C, D, X, Y, и RT.
Семейство A
Семейство A включает в себя репликативные и восстановительные ДНК-полимеразы. Репликативные члены этого семейства представлены, например, хорошо исследованной ДНК-полимеразой вируса Т7 или эукариотической митохондриальной ДНК-полимеразой γ. Среди восстановительных полимераз мы находим такие примеры как ДНК-полимераза I E. coli, полимераза I из Thermus aquaticus или полимераза I Bacillus stearothermophilus. Восстановительные полимеразы участвуют в процессе устранения ошибок в собираемой ДНК, а также в обработке фрагментов Оказаки.
Семейство B
В семейство B в основном входят восстановительные полимеразы, в том числе основные эукариотические ДНК-полимеразы α, δ, и ε, а также ДНК-полимераза ζ. К этому семейству также относят ДНК-полимеразы некоторых бактерий и бактериофагов, например бактериофагов T4, Phi29 и RB69. Эти ферменты используются в синтезе и 3’-5’, и 5’-3’ моноцепей ДНК. Отличительной особенностью полимераз этого семейства является замечательная точность репликации. Многие также обладают сильным 3’-5’-экзонуклеазным действием (за исключением ДНК-полимераз α и ζ, у которых способности корректировать ошибки не наблюдается).
Семейство C
Полимеразы этого семейства — в основном бактериальные хромосомные репликативные ферменты, обладающие, кроме того, 3’-5’-экзонуклеазным действием.
Семейство D
Полимеразы этого семейства недостаточно изучены. Все известные образцы считаются репликативными полимеразами и обнаружены у архей субдомена Euryarchaeota.
Семейство X
К семейству Х относится широко известная эукариотическая ДНК-полимераза β, а также и другие, такие как σ, λ, μ и концевая дезоксинуклеотидил-трансфераза (TdT). ДНК-полимераза β необходима для осуществления процесса восстановления поврежденных участков ДНК. Полимеразы λ и μ участвуют в негомологическом соединении — процессе восстановления разрывов двойной спирали.. TdT экспрессируется только в лимфоидной ткани и добавляет «n нуклеотидов» к разрывам двойной спирали, образующимся во время В(Р)С-рекомбинации. Дрожжи Saccharomyces cerevisiae обладают лишь одной полимеразой X, Pol4, участвующей в негомологическом соединении.
Семейство Y
Полимеразы этого семейства отличаются от прочих низкой производительностью на целостных шаблонах, а также способностью осуществлять репликацию на шаблонах поврежденных ДНК. Вследствие этого члены семейства называются полимеразами транслезионного синтеза. В зависимости от характера повреждения (лезии) ТЛС-полимеразы могут восстановить исходную цепочку. Ошибка может и не быть восстановлена, что приводит к мутациям. Страдающие Xeroderma pigmentosum, например, обладают мутантным геном ДНК-полимеразы η (eta), который толерантен к повреждениям, однако другие полимеразы, например ζ (относящаяся к семейству B), страдают от мутаций, что, как считается, приводит к предрасположенности к онкологическим заболеваниям.
Другие члены этого семейства — человеческие полимеразы ι, κ, а также концевая дезоксинуклеотидил-трансфераза Rev1. У E.coli имеются две ТЛС-полимеразы: IV (DINB) и V (UMUC).
Семейство RT
Основная статья: Обратная транскриптаза
Семейство обратных транскриптаз (название семейства происходит от англ. reverse transcriptase) содержит полимеразы, обнаруженные как у ретровирусов, так и у эукариот. Они являются РНК-зависимыми ДНК-полимеразами, то есть, в отличие от описанных выше ферментов, используют в качестве матрицы для синтеза РНК, а не ДНК. Эукариотические обратные транскриптазы в основном представлены теломеразами. Эти полимеразы используют шаблонную РНК для синтеза цепочки ДНК.
Уровни структурной организации
Изогнутая как спираль полинуклеотидная цепь – это первичная структура, которая имеет определенный качественный и количественный набор мононуклеотидов, связанных 3’,5’-фосфодиэфирной связью. Таким образом, каждая из цепей имеет 3’-конец (дезоксирибоза) и 5’-конец (фосфатный). Участки, которые содержат в себе генетическую информацию, названы структурными генами.
Двухспиральная молекула – это вторичная структура. Причем ее полинуклеотидные цепи антипараллельны и связываются водородными связями между комплементарными основаниями цепей. Установлено, что в каждом витке этой спирали содержится 10 нуклеотидных остатков, длина ее равняется 3,4 нм. Эту структуру поддерживают также Ван-дер-Ваальсовы силы взаимодействия, которые наблюдаются между основаниями одной цепи, включающие отталкивающие и притягивающие компоненты. Эти силы объясняются взаимодействием электронов в соседних атомах. Электростатическое взаимодействие также стабилизирует вторичную структуру. Оно возникает между заряженными положительно молекулами гистонов и заряженной отрицательно нитью ДНК.
Третичная структура – это намотка цепей ДНК на гистоны или суперспирализация. Описано пять видов гистонов: Н1, Н2А, Н2В, Н3, Н4.
Укладка нуклеосом в хроматин – это четвертичная структура, поэтому молекула ДНК, имеющая длину несколько сантиметров, может складываться до 5 нм.
Надежда на быстрый успех
О нормальной — то есть довольно халатной — работе этого фермента и механизмах закрепления его ошибок в геноме очень доходчиво рассказывает статья «Следы полимеразы α» . — Ред.
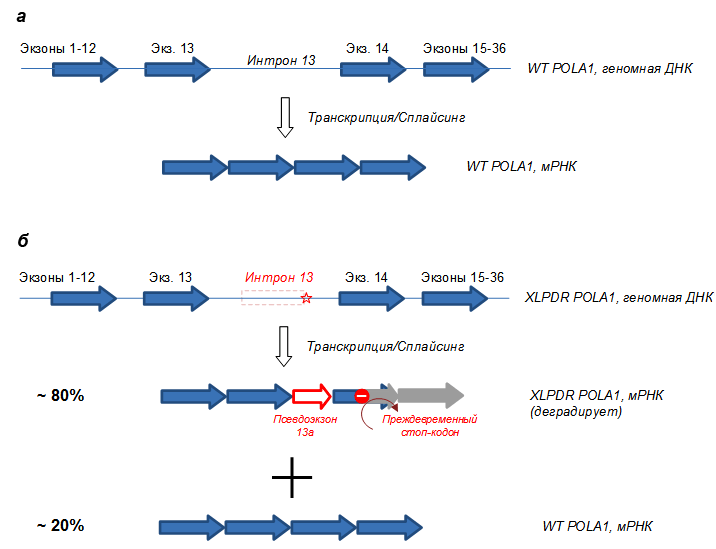
Рисунок 2. Схема гена POLA1 и мРНК у здоровых людей (а) и у больных XLPDR (б). В результате точечной замены в интроне 13 (звездочка) образуется ложный альтернативный сайт сплайсинга, и бόльшая часть пре-мРНК образует нежизнеспособную мРНК, которая быстро деградирует. Меньшая часть мРНК всё же собирается правильно, что обеспечивает выживание клеток (репликацию), однако этого количества POLA1 не хватает для выполнения альтернативных функций (см. дальше).
Структура двойной спирали ДНК
Предпочтительное направление изгиба ДНК определяется стабильностью укладки каждого основания поверх следующего. Если на одной стороне спирали ДНК всегда находятся неустойчивые этапы штабелирования основы, то ДНК предпочтительно будет отгибаться от этого направления. Соединение двух цепей ДНК в спираль осуществляют молекулы, зависящие от этого направления. По мере увеличения угла изгиба они играют роль стерических препятствий, проявляя способность прокатывать остатки по отношению друг к другу, особенно в малой канавке. Отложения A и T будут предпочтительно встречаться в небольших канавках внутри изгибов. Этот эффект особенно проявляется в связывании ДНК-белка, когда индуцируется жесткий изгиб ДНК, например, в нуклеосомных частицах.
Молекулы ДНК с исключительным изгибом могут стать изгибающимися. Это было впервые обнаружено в ДНК трипаносоматида кинетопласта. Типичные последовательности, которые вызывают это, содержат отрезки 4-6 T и A, разделенные по принципу G и C, которые содержат остатки A и T в фазе с малой канавкой на одной стороне молекулы.
Внутренняя изогнутая структура индуцируется «прокруткой винта» пар оснований друг относительно друга, что позволяет создавать необычные бифурцированные водородные связи между базовыми ступенями. При более высоких температурах эта структура денатурирована, и поэтому собственный изгиб теряется.
Вся ДНК, которая изгибается анизотропно, имеет, в среднем, более длительный упор и большую осевую жесткость. Эта повышенная жесткость необходима для предотвращения случайного изгиба, который заставит молекулу действовать изотропно.
Кольцевание ДНК зависит как от осевой (изгибной) жесткости, так и от крутильной (вращательной) жесткости молекулы. Чтобы молекула ДНК успешно циркулировала, она должна быть достаточно длинной, чтобы легко сгибаться в полный круг и иметь правильное количество оснований, чтобы концы находились в правильном вращении, дабы обеспечить возможность склеивания спиралей. Оптимальная длина для циркулирования ДНК составляет около 400 пар оснований (136 нм). Наличие нечетного числа поворотов представляет собой значительный энергетический барьер для кругооборотов, например, молекула пары 10,4 х 30 = 312 будет циркулировать в сотни раз быстрее, чем 10,4 х 30,5 ≈ 317-молекулы.
Вам будет интересно:Командор — это: значение слова и синонимы
Немного теории для не биологов
Для понимания предмета и самой сути доказательства роли ДНК в наследственности напомним лишь некоторые общие понятия и термины, применяемые в тексте. Молекулярные биологи и другие профессиональные биологи могут эту часть не читать – понятия даются в упрощенном варианте для интересующейся части читателей. Хотя современная специализация в биологии сегодня настолько возросла, что профессионал-эколог не всегда прорвется к пониманию сути эволюционных механизмов, а ботанику совсем не однозначно понятна специфика развития эмбриона лягушки. Итак, вот эти термины:
- ДНК (дезоксирибонуклеиновая кислота) и РНК (рибонуклеиновая кислота) – длинные и большие молекулы, состоящие из мономеров – нуклеотидов.
- ДНК и РНК называют нуклеиновыми кислотами.
- ДНК и РНК образованы всего четырьмя нуклеотидами (три одинаковых, один разный в ДНК и РНК) – нуклеотиды универсальны для всего живого на планете. Это сложные органические соединения из азотистого основания, углеводного остатка и фосфорной кислоты. Называются они адеин, гуанин, тимин и цитозин (урацил).
- Нуклеотиды образуют триплеты – они кодируют одну аминокислоту из двадцати.
- Триплеты образуют цепочки в составе нуклеиновых кислот, что соответствует цепочке аминокислот или одному специфическому белку. Белки – основа жизни на планете, они специфичны и уникальны.
- Ген – это кусочек нуклеиновой кислоты, отвечающий за один белок.
- Геном – совокупность всего генетического материала организма.
Post Scriptum
К сожалению, в России подобные исследования находятся в зачаточном состоянии. Только несколько лабораторий пытаются работать с отечественным палеонтологическим и археологическим материалом, который подобно углеводородам уходит за рубеж. В 2014 году сотрудники Центра «Биоинженерия» РАН и НИЦ «Курчатовский институт» впервые в России отсеквенировали последовательность митохондриальной ДНК (и определили её гаплогруппу) у древнего человека, обитавшего на территории Северного Кавказа в эпоху ранней бронзы . Эта пионерская для России работа, возможно, даст толчок отечественной археологической и палеогенетике.