Археоптерикс
Содержание:
Упоминание в книгах
- В романе Майкла Крайтона «Парк юрского периода» рассказывается о воссоздании Тираннозавров в лаборатории InGen. Пробелы в геноме заполняют ДНК птиц. Полученные клоны не похожи на исторических предшественников. Они могут рычать и плавать. У них длинный и подвижный язык. Но их кожа чувствительна к ультрафиолету, поэтому динозавры предпочитают держаться в тени. Шкура клонов красно-коричневая.
- Во второй книге «Затерянный мир» пара Тираннозавров обитает на острове Сорна и высиживает яйца в гнезде. Команда Льюиса Доджсона пытается украсть яйца, но план провалился. В результате гибнет Джордж Бейлизтон. Эдди Кар подбирает раненого детёныша Тираннозавра и несёт в прицеп. Команда накладывает гипс на сломанную лапку животного, но вернуть его не успевает. Родители нашли трейлер, и как только маленький Тираннозавр оказывается в безопасности, пара динозавров атакует прицепы и сбрасывает их со скалы.
Энциклопедия в дополненной реальности «Динозавры:от компсогната до рамфоринха»
Размножение Тираннозавров
На видео представлена одна из реконструкций секса тираннозавров.
Из-за крупных размеров животным приходилось прилагать усилия для совершения полового акта. Самцы тираннозавров были чуть меньше самок. Во время спаривания самец забирался или прижимался к лежащей или стоящей самке. Самцы могли иметь копулятивный орган, который помогал ему дотянуться до самки. Оплодотворение яйцеклеток проходило внутри самки. Во время беременности вокруг яйцеклеток формировалась скорлупа. Спустя несколько месяцев самка откладывала яйца в гнездо, а через несколько недель потомство тираннозавров появлялось на свет.
Передвижение
 На видео — бег тираннозавра в замедленном варианте
На видео — бег тираннозавра в замедленном варианте
Эти динозавры перемещались на задних лапах, поочерёдно переставляя ноги. Удерживать равновесие хищнику помогал длинный тяжёлый хвост. Голову ящер держал прямо. Походку тираннозавра удалось воссоздать благодаря изучению следов. Когда динозавр стоял, задние конечности располагались параллельно друг другу. Бёдра принимали почти вертикальное положение, но при ходьбе переносились к серединной линии тела ящера.
Ноги животного двигались вперёд-назад и из стороны в сторону. Эта особенность движения отчётливо заметна по следам, оставленным хищником во время бега.
Description
Archaeopteryx lived in the early Jurassic Period, 150.8–148.5 million years ago. Most of the specimens of Archaeopteryx that have been found come from the Solnhofen limestone in Bavaria, Germany, which is a lagerstätte, a rare and stunning geological formation known for its superbly full fossils.
Archaeopteryx was close to the size of a raven, with broad wings that were rounded at the ends and a long tail. It could reach up to 20 inches in length, with a weight of 1.8 to 2.2 pounds.Archaeopteryx feathers, though less documented than its other traits, looked a lot like bird feathers in structure and design. But,Archaeopteryx had a lot of theropod dinosaur traits too. Unlike modern birds, Archaeopteryx had small teeth as well as a long bony tail, traits which Archaeopteryx shared with other dinosaurs of the time.
As it shows quite a few traits of both birds and dinosaurs, Archaeopteryx has often been deemed a link between them. In the 1970s, John Ostrom, following T. H. Huxley’s lead in 1868, argued that birds evolved within theropod dinosaurs and Archaeopteryx was a critical piece of evidence for this argument; it had a number of bird traits, such as a wishbone, flight feathers, wings and a partially reversed first toe, and a number of dinosaur and theropod features. For instance, it has a long ascending process of the ankle bone, interdental plate, an obturator process of the ischium, and long chevrons in the tail. In particular, Ostrom found that Archaeopteryx was remarkably similar to the theropod family Dromaeosauridae.
The first bones of Archaeopteryx were found in 1861; just two years after Charles Darwin wrote On the Origin of Species. Archaeopteryx seemed to prove Darwin’s theories and has since been a key piece of proof for the origin of birds, the transitional fossil debate, and proof of evolution. Indeed, more research on dinosaurs from the Gobi Desert and China has since given more proof of a link between Archaeopteryx and the dinosaurs, such as the Chinese feathered dinosaurs. Archaeopteryx is close to the ancestry of modern birds, and it shows most of the features one would expect in an ancestral bird. But, it may not be the direct ancestor of living birds, and it is not clear how much evolutionary divergence was already found in other birds at the time.
Notes/Trivia[edit | edit source]
For information pertaining specifically to the real-world Archaeopteryx, see the relevant Wikipedia article.
- Dossier revealed on January 4, 2016
- Despite being a winged creature, it can be carried by other (rideable) flying creatures.
- On Scorched Earth, Archaeopteryx can be found wandering the cactus forests.
- On Scorched Earth, there is a humorous glitch that can happen on rare occasions regarding Archaeopteryx. They can be seen flying vertically into the sky for seemingly an endless amount of time, soaring higher and higher on a slow, 90-degree climb towards the top of the map’s skybox. This might be a glitch if an Archaeopteryx has trouble latching onto a Joshua tree, as Archaeopteryx can also be found humorously flying circles around a single tree while calling repetitively.
- Falling with an Archaeopteryx can still sometimes be dangerous; the game uses vectors to determine your flight paths, which means that a large amount of speed down vertically can take time to correct horizontally.
- Though the Archaeopteryx has often been shown a an example of a missing link between dinosaurs and birds, it is incorrect to assume that the Archaeopteryx was an ancestor to birds. Instead, it was a part of the early radiation of Avialae(animals more closely related to a modern bird, like the chicken, than to other memebers of the Paraves, like the Deinonychus) which included the ancestor of Aves(modern birds) in the Late Jurassic.
- Humorously, ever since v279.22, or TLC Phase 2, came to PC, and the Raptor got the pounce ability, there has been a slight glitch. When a survivor gets pounced while holding an Archaeopteryx, ocassionally it too will display the animation as if it were being pouced on its own but still being held. Being pounced just one time while holding the Archa can also tweak the angle at which it faces, meaning while you glide with it in one direction, the Archa will appear to be flying at an angle. This is fixed when the Archa is dropped and picked up again.
- Real world Archaeopteryx were carnivores that ate small animals and insects, however Ark’s Archaeopteryx have adapted to only eating chitin.
- The Archaeopteryx has a curious and stark resemblance to the Microraptor, the only significant difference between the 2 being the Microraptor’s head crest and incredible speed.
- On Valguero, the Archaeopteryx spawns in the Southeastern half of the Redwoods zone.
- Strangely enough on the Ocean Biome in Genesis, Archaeopteryx does not run away from the survivors even if they are touching it.
In other languages
| Archaeopteryx | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
| Fossils | |||||
|---|---|---|---|---|---|
|
|
|||||
|
Описание
Размер подростковой особи голотипа (синий) и взрослого образца (зелёный) в позициях стойки и полёта, в сравнении с человеком
Птеродактиль известен по более чем 30 ископаемым образцам и, хотя большинство из них являются подростковыми особями, многие сохранили полные скелеты. Pterodactylus antiquus был небольшим птерозавром, размах крыльев взрослой особи оценивается в 1 м (единственный взрослый экземпляр представлен с отделённым черепом). Предполагается, что размеры прочих видов были меньше. Возможно, некоторые из них представляют собой неполовозрелых особей птеродактиля, а также других птерозавров, живших в то же время, таких, как ктенохазма, германодактиль, Aerodactylus, Aurorazhdarcho и Gnathosaurus.
Черепа взрослых птеродактилей были длинными и тонкими и несли около 90 узких конических зубов. Зубы были длинными на кончиках обеих челюстей и становились тем меньше, чем глубже в пасти находились. Это отличало птеродактилей от родственных видов, чьи зубы отсутствовали на кончике верхней челюсти и были одинаковыми по размеру. Зубы птеродактиля также тянулись в глубь пасти гораздо дальше, чем у близкородственных видов, поскольку некоторые зубы присутствуют даже в передней части предглазнично-носового окна, крупнейшего отверстия в черепе. В отличие от родственных видов, череп и челюсти птеродактиля были прямыми, а не загнутыми вверх.
Птеродактиль в представлении художника
Птеродактиль, подобно родственным видам, имел гребень на черепе, который состоял, в основном, из мягких тканей. У взрослых особей гребень проходил между задней кромкой предглазничного окна и затылочной частью черепа. По крайней мере, у одного ископаемого образца гребень имел короткое костное основание, также находимое у родственных птерозавров, таких, как германодактиль. Костные гребни были найдены только на больших скелетах взрослых особей птеродактилей, и это указывает на то, что данная структура выполняла демонстрационную функцию и становилась больше по мере роста животного. Беннет (2013) отмечает: другие авторы утверждают, что гребень из мягких тканей у птеродактиля продолжался и за черепом; сам же Беннет не нашёл никаких доказательств этому. Два образца P. antiquus (голотип BSP AS I 739 и неполный череп BMMS 7 — самый большой череп P. antiquus) несут низкий костный гребень на черепах; у BMMS 7 он 47,5 мм длиной (примерно 24 % от расчётной общей длины черепа) и имеет максимальную высоту 0,9 мм над глазницей.
Paleobiology
Plumage
Specimens of Archaeopteryx were best known for their well-developed flight feathers. They were crooked and showed the form of flight feathers in birds, with stable vanes. The tail feathers were less crooked and had firm vanes, just like in birds, but the thumb did not yet bear a separately movable tuft of stiff feathers.
The body plumage of Archaeopteryx is less well documented and has just been properly researched in the well-preserved Berlin specimen. Thus, as more than one species seems to be involved, the research of the Berlin specimen’s feathers does not necessarily hold true for the rest of the species of Archaeopteryx. In the Berlin specimen, there are «trousers» of well-developed feathers on the legs; some of which seem to have a plain form feather structure but are quite decomposed.But, in part they are firm and thus can support flight.
A patch of feathers ran down the back which looked much like those of birds in that they were aligned and firm, though not as stiff as the flight-related feathers. Apart from that, the feather traces in the Berlin specimen are limited to a sort of «proto-down» like that found in the dinosaur Sinosauropteryx, decomposed and soft, and might have looked more like fur than like feathers in life (though not in their microscopic structure). These are on the rest of the body, as far as such structures are both preserved and not wiped out by preparation, and the lower neck.
But, there is no sign of feathers on the upper neck and head. While these may have been nude, this may still be an artifact of preservation. It seems that most Archaeopteryx specimens became embedded in anoxic sediment after drifting some time on their back in the sea—the head and neck and the tail are for the most part bent down, which may mean that the specimens had just started to rot when they were embedded, with tendons and muscle relaxing so that the characteristic shape of the fossil specimens was gained. This would mean that the skin was already softened and loose, which is helped by the fact that in some specimens the flight feathers were starting to come off at the point of embedding in the sediment. So it is thought that the pertinent specimens moved on the edge the sea bed in shallow water for some time till buried, the head and upper neck feathers sloughed off, while the more firmly attached tail feathers remained.
Color
In 2011, graduate student Ryan Carney and colleagues did the first colour study on an Archaeopteryx specimen. The team found the structure of melanosome in the single-feather specimen described in 1861. The resultant structure was then compared to that of 87 bird species and was shown to very likely be black. The feather studied was most likely one covert, which would have covered part of the main feathers on the wings. While the study does not mean that Archaeopteryx was all black, it does hint that it had some black color such as the coverts. Carney pointed out that this is in line with what we know of modern flight traits, in that black melanosomes have structural properties that make feathers strong for flight.
Примеры среди животных
Древнейшими представителями земноводных считают ихтиостеги. Их считают переходным звеном между кистеперых рыбами и земноводными. Несмотря на то, что в ихтиостеги была пятипалая концовка, адаптированная к жизни на суше, значительную часть жизни они проводили как рыбы, имели хвостовой плавник, боковую линию и некоторые другие признаки рыб.
Батрахозавры, существовавшие в каменноугольный и пермский периоды, рассматривают как переходную форму между земноводными и пресмыкающимися. Батрахозавры, хоть и проводили жизнь во взрослой стадии на суше (подобно пресмыкающихся), были тесно связаны с водоемами и сохранили ряд признаков, присущих земноводным, в частности, откладывания икры и развитие личинок в воде, наличие жабр и тому подобное.
Обнаружено большое количество пресмыкающихся, которые выработали способность летать, часть из них имела перья, поэтому их рассматривают как переходные формы между пресмыкающимися и птицами. Наиболее известен археоптерикс. Он был размером примерно с современную ворону. Формой тела, строением конечностей и наличием оперения подобный современных птиц, возможно, летал. Общим с пресмыкающимися была особое строение таза и ребер, наличие клюва с коническими зубами, по три свободные пальцы на крыльях, подвийноувигнути позвонки, длинный хвост с 20-21 позвонка, кости могло не пневматизовани, грудная кость без киля. Другие известные переходные формы между пресмыкающимися и птицами — протоавис, конфуциусорниса.
Большое количество ископаемых форм звероподобных пресмыкающихся (синапсиды, терапсид, пеликозавров, различных динозавровых и др.), Найденных во многих районах земного шара, существовали в юрский и меловой периоды, сочетающие признаки пресмыкающихся и млекопитающих, раскрывают возможные направления и способы становления различных групп четвероногих , в частности млекопитающих. Например, звероподобный пресмыкающееся из группы терапсид — лиценопс (Lycaenops) по развитию костей ротовой полости, дифференцированием зубов на клыки, резцы, по резцовые зубы и рядом других признаков строения тела напоминает хищных млекопитающих, хотя по другим признакам и образом жизни это были настоящие пресмыкающиеся.
Одной из форм, сохранилась в ископаемом состоянии является амбулоцетус Ambulocetus natans («ходячий кит») — переходная форма между наземными млекопитающими и китообразными, которые являются вторинноводнимы формами. Внешне животное напоминало нечто среднее между крокодилом и дельфином. Кожа должна частично редуцированную шерсть. Животное имело лапы с перепонками; хвост и конечности приспособлены как вспомогательные органы передвижения в воде.
Спор о постепенности эволюционных преобразований
Переходные формы являются объектом исследований в споре между градуалистами и пунктуалистами. Градуалисты считают, что эволюция является постепенным, непрерывным процессом. Пунктуалисты, или сторонники теории прерывистого равновесия считают, что виды в течение долгого времени остаются неизменными, а изменения происходят в очень короткий промежуток времени. Переходные формы являются аргументом обеих сторон. Пунктуалисты утверждают, что большинство видов появляются внезапно и на протяжении всей своей истории меняются слабо. Градуалисты отмечают, что среди позвоночных нет ни одного бесспорного примера очень быстрого перехода от одного вида к другому, большинство переходов вообще плохо задокументированы ископаемыми. Также много хорошо описанных внутривидовых изменений. Например, описаны изменения среднекайнозойских кроликов, миоценовых грызунов, гиппарионов, эоценовых парнокопытных, переход между родами эоценовых приматов.
Taxonomy
The relationships of the specimens are problematic. Most specimens have been given their own species at one point or another. The Berlin specimen has been referred to Archaeornis siemensii, the Eichstätt specimen to Jurapteryx recurva, the Munich specimen to Archaeopteryx bavarica and the Solnhofen specimen was referred to Wellnhoferia grandis.
Recently, it has been argued that all the specimens belong to the same species (, 17 April 2004, p.17). However, significant differences exist among the specimens. In particular, the Munich, Eichstätt, Solnhofen and Thermopolis specimens differ from the London, Berlin, and Haarlem specimens in being smaller or much larger, having different finger proportions, having more slender snouts, lined with forward-pointing teeth and possible presence of a sternum. These differences are as large as or larger than the differences seen today between adults of different bird species. However, it is also possible that these differences could be explained by different ages of the living birds.
Способность к полету
Строение перьевого покрова (ассиметричное строение маховых перьев) подтверждает тот факт, что археоптерикс был способен летать. Но ученые еще не пришли к общему мнению о том был ли это полет примитивным (планирующим), или более сложным (машущим), как у поздних птиц.
При оценке нагрузки на крыло ученые утверждают, что этот древний ящер был способен только к планирующему полету, так как не мог поднять крыло выше уровня спины, что является необходимым условием для замаха.
Другие исследования указывают что форма тела и размер крыла были нетипичны для планирующих птиц, а вилочка и коракоид служили для прикрепления мышц, двигающих крыло.
Интересным представляется гипотеза, что археоптерикс мог совершать небольшие движения крыльями без замаха. Эти движения совершались во время планирующего полета.
Строение скелета
В большинстве работ палеонтологов археоптерикс описывается как первая птица произошедшая от ящеротазовых динозавров. Некоторые исследователи считают его потомков тероподов. Но на самом деле точное систематическое положение существа до конца не определено, есть версия, что археоптерикс — переходная форма между рептилиями и птицами. Причиной двойственной принадлежности вида к древним пресмыкающимся и птицам стало наличие анатомических особенностей, характерных для каждой из групп.
Строение позвоночника и верхних конечностей
У археоптерикса передние конечности были длинными, с тремя хорошо развитыми пальцами. Формула фаланг: 2–3–4. Заканчивались пальцы длинными загнутыми когтями. Кости пясти и запястья не срослись, полулунная кость составляла часть запястья.
Шейные позвонки археоптерикса двояковогнуты: вогнуты сзади и спереди, что присуще динозаврам семейства дейнонихозавров.
В крестовом отделе позвоночника у археоптерикса пять несросшихся позвонков (у птиц часть поясничных, крестцовые и начальные хвостовые позвонки срастаются в одну кость — сложный крестец, срастаются тазовые кости, а оставшиеся хвостовые позвонки срастаются, образую кость — пигостиль). Подвздошная, седалищная и лобковая кости археоптерикса не образуют единую кость, дистальные концы лобковых костей соединяются и образуют лобковый симфиз, а хвост у археоптерикса длинный и состоит из 21–23 отдельных позвонков.
У археоптерикса были найдены вентральные ребра (гастралии), что характерно для пресмыкающихся, без крючковидных отростков (эти отростки у птиц обеспечивали прочность грудной клетки). Ключицы археоптерикса срастались и образовывали вилочку, что характерно как для птиц, так и для динозавров.
Строение нижних конечностей
Голень состоит из большой и малой берцовых костей, примерно одинаковой длины. Часть костей предплюсны и плюсны не срослись, а это значит, что у археоптерикса отсутствовала цевка.
Большой палец противопоставлен остальным, что является характерным для птиц . На втором, самом длинном пальце задней конечности, рос длинный коготь, что присуще динозавром семейства дейнонихозавров.
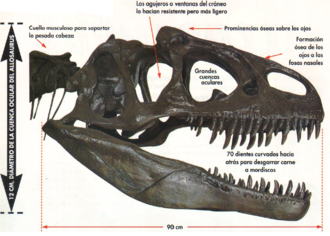
Строение черепа
На предчелюстных, челюстных и зубных костях археоптерикса росли мелкие конические зубы, как у современных крокодилов. Предчелюстные кости не срастались. У нижней и верхней челюсти отсутствовал роговой чехол, а значит клювом археоптерикс не обладал. На затылке, сзади, у археоптерикса расположено отверстие, соединяющее позвоночный канал и череп.
Мозг археоптерикса такого же объема, что у современных птиц и в три раза больше, чем у рептилий того же размера. Полушария мозга археоптерикса не окружают обонятельные трактаты и они пропорционально меньше, чем у современных птиц.
Мозжечок динозавра больше, чем у тероподов, но меньше, чем у современных птиц. Зрительные доли мозга расположены спереди. Полет археоптерикса был невозможен без увеличенных зрительных долей и мозжечка.
Положение переднего и бокового полукружных каналов у археоптерикса типично для архозавров, но передний канал длиннее и изогнут в обратном направлении.
Перья археоптерикса
Именно это окаменевшее перо археоптерикса с которого началась история пернатых динозавров
Лондонский и берлинский экземпляры археоптерикса сохранили отпечатки перьевого покрова. Оперение — это ключевая особенность, по которой археоптерикса относят к птицам.
Перья археоптерикса, как и у современных птиц, подразделялись на маховые, рулевые и контурные. Маховые и рулевые перья археоптерикса имеют стержень пера, бородки 1-го порядка и бородки 2-го порядка. Для маховых перьев характерно ассиметричное строение опахал, что типично для современных птиц, способных к полёту. Отличие перьевого покрова современных птиц от археоптерикса — наличие крылышка (подвижного пучка перьев на большом пальце передних конечностей).