Химический синапс
Содержание:
Компоненты
Типы
Эта система работает таким образом, что тормозные постсинаптические потенциалы суммируются во времени с подпороговыми или надпороговыми возбуждающими потенциалами, в результате чего уменьшается результирующий постсинаптический потенциал. Эквивалентные по модулю возбуждающие (положительные) и тормозные (отрицательные) постсинаптические потенциалы дают в сумме нейтральное состояние, взаимно отменяя действие друг друга на клетку. Баланс между возбуждающими и тормозными постсинаптическими потенциалами очень важен для интеграции клеткой всей поступающей от различных возбуждающих и тормозных синапсов электрической и химической информации.
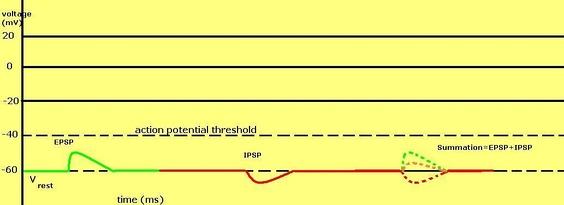
Схема, показывающая воздействие одновременно возбуждающего и тормозного потенциалов на нейрон и их суммацию: в данном конкретном случае сумма потенциалов оказывается всё равно ниже порога возникновения потенциала действия.
Дополнительные факторы
Размер нейрона также может влиять на эффект, который оказывает на клетку тормозной постсинаптический потенциал. Простое и мгновенное временное суммирование постсинаптических потенциалов имеет место быть в нейронах сравнительно небольшого размера, тогда как в крупных нейронах большее количество синапсов, метаботропных и ионотропных рецепторов, а также наличие длинных аксонов и большее расстояние от синапсов до тела нейрона позволяет нейронам ещё некоторое время продолжать электрическое и химическое общение с другими нейронами (то есть пребывать в состоянии возбуждения), несмотря на наличие тормозных потенциалов на удалённых от тела синапсах, пока тормозящий сигнал «путешествует» до тела клетки.
Модель пластичности, зависимая от времени спайка
Модель пластичности, зависимая от времени спайка (spike-timing-dependent plasticity, STDP) это вид синаптической пластичности, который регулирует силу связей в зависимости от относительного времени между активацией нейрона и входными потенциалами действия (спайками на входе). Согласно этой модели пластичности, если входной спайк следует в среднем непосредственно перед активацией нейрона, то такая связь усиливается, а если следует в среднем непосредственно после активации нейрона, то такая связь ослабляется. Таким образом усиливается влияние входов, которые могут служить причиной активации нейрона, и ослабляется влияние входов, сигналы которых не могли и не могут быть причиной активации. Данный вид пластичности хорошо согласуется с принципом Хебба для обучения искусственных нейронных сетей, согласно которому должна усиливаться связь между узлами, активирующимися одновременно. Так как в биологической системе сигналы распространяются не мгновенно, а с задержкой, то чтобы активация одного нейрона могла быть причиной активации другого, первый должен активироваться немного раньше.
Классификации синапсов
Основные элементы электрического синапса (эфапса): а — коннексон в закрытом состоянии; b — коннексон в открытом состоянии; с — коннексон, встроенный в мембрану; d — мономер коннексина, е — плазматическая мембрана; f — межклеточное пространство; g — промежуток в 2-4 нанометра в электрическом синапсе; h — гидрофильный канал коннексона
По механизму передачи нервного импульса
- химический — это место близкого прилегания двух нервных клеток, для передачи нервного импульса через которое клетка-источник выпускает в межклеточное пространство особое вещество, нейромедиатор, присутствие которого в синаптической щели возбуждает или затормаживает клетку-приёмник.
- электрический (эфапс) — место более близкого прилегания пары клеток, где их мембраны соединяются с помощью особых белковых образований — коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе — 3,5 нм (обычное межклеточное — 20 нм). Так как сопротивление внеклеточной жидкости мало (в данном случае), импульсы через синапс проходят не задерживаясь. Электрические синапсы обычно бывают возбуждающими.
- смешанные синапсы — пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Наиболее распространены химические синапсы. Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические.
По местоположению и принадлежности структурам
- периферические
- нервно-мышечные
- нейросекреторные (аксо-вазальные)
- рецепторно-нейрональные
- центральные
-
аксо-дендритические — с дендритами
аксо-шипиковые — с дендритными шипиками, выростами на дендритах;
, в том числе
- аксо-соматические — с телами нейронов;
- аксо-аксональные — между аксонами;
- дендро-дендритические — между дендритами;
-
аксо-дендритические — с дендритами
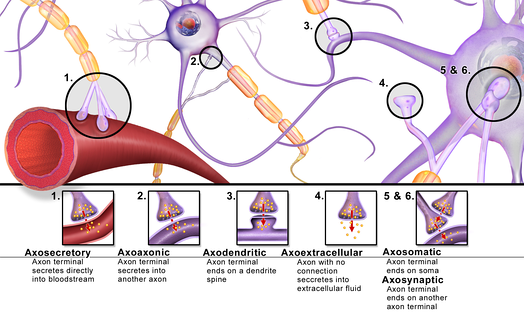
Различные варианты расположения химических синапсов
По нейромедиатору
- аминергические, содержащие биогенные амины (например, серотонин, дофамин
в том числе адренергические, содержащие адреналин или норадреналин;
);
- холинергические, содержащие ацетилхолин;
- пуринергические, содержащие пурины;
- пептидергические, содержащие пептиды.
При этом в синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора.
По знаку действия синапса
- возбуждающие
- тормозные.
Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембраны, которая может вызвать потенциал действия при определённых условиях), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор — глицин) и ГАМК-ергические синапсы (медиатор — гамма-аминомасляная кислота).
Тормозные синапсы бывают двух видов:
1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала;
2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение.
В некоторых синапсах присутствует постсинаптическое уплотнение — электронно-плотная зона, состоящая из белков. По её наличию или отсутствию выделяют синапсы асимметричные и симметричные. Известно, что все глутаматергические синапсы асимметричны, а ГАМКергические — симметричны.
В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы.
К специальным формам синапсов относятся шипиковые аппараты, в которых с синаптическим расширением контактируют короткие одиночные или множественные выпячивания постсинаптической мембраны дендрита. Шипиковые аппараты значительно увеличивают количество синаптических контактов на нейроне и, следовательно, количество перерабатываемой информации. «Не-шипиковые» синапсы называются «сидячими». Например, сидячими являются все ГАМК-ергические синапсы.
Ингибирующие рецепторы
Существует два типа ингибирующих рецепторов:
Ионотропные рецепторы
Ионотропные рецепторы (также известные как открываемые лигандом ионные каналы) играют важную роль в быстрой генерации тормозных постсинаптических потенциалов. Нейромедиатор связывается со специфическим доменом рецептора — так называемым сайтом или доменом связывания лиганда (ligand-binding site or domain) рецептора, расположенным на наружной стороне поверхностной мембраны клетки (обращённым в синаптическую щель). Это приводит к изменению пространственной конфигурации рецептора и к открытию в нём ионного канала, который образуется внутри эндомебранного (проходящего сквозь мембрану) домена рецептора. В результате возникает быстрый входящий или исходящий ток ионов — внутрь или наружу клетки. Ионотропные рецепторы способны продуцировать очень быстрые изменения постсинаптического потенциала — в течение миллисекунд после генерации потенциала пресинаптической клеткой. Ионные каналы способны влиять на амплитуду и временные характеристики потенциала действия клетки в целом. Ионотропные ГАМК-рецепторы, сопряженные с каналами ионов хлора, являются мишенью действия многих лекарств, в частности барбитуратов, бензодиазепинов, ГАМК-аналогов и агонистов, ГАМК-антагонистов, таких, как пикротоксин. Алкоголь также модулирует ионотропные ГАМК-рецепторы.
Метаботропные рецепторы
Метаботропные рецепторы, большинство из которых относятся к семейству G-белок-связанных рецепторов, не содержат встроенных в их структуру ионных каналов. Вместо этого они содержат внеклеточный домен связывания лиганда (ligand-binding domain) и внутриклеточный домен связывания с первичным эффекторным белком, которым наиболее часто бывает G-белок. Связывание агониста с метаботропным рецептором приводит к такому изменению конфигурации рецептора, при котором активируется первичный эффекторный белок. Так, например, в случае G-белка активация связанного с ним рецептора приводит к диссоциации β- и γ- субъединиц G-белка в виде βγ-димера и к активации ими ряда «добавочных» внутриклеточных сигнальных путей (в частности en:GIRK), в то время как активированная α-субъединица G-белка изменяет активность классического аденилатциклазного пути (повышает в случае стимулирующего Gs белка и угнетает в случае ингибирующего Gi). Это, в свою очередь, приводит к изменению внутриклеточной концентрации вторичного посредника — циклического АМФ — повышению, в случае повышения активности аденилатциклазы, или снижению, в случае снижения. А изменение концентрации циклического АМФ влияет на активность цАМФ-зависимой протеинкиназы A — вторичного эффектора. Повышение или снижение активности протеинкиназы А запускает нисходящий эффекторный каскад вплоть до эффекторов N-го порядка. В частности, открываются или закрываются ионные каналы.
Ингибирующие метаботропные рецепторы связаны всегда с ингибирующим подтипом G-белка, то есть с Gi. Таким образом, они угнетают активность аденилатциклазы и снижают концентрацию циклического АМФ, эффективно угнетая тем самым активность протеинкиназы А. Помимо этого, они активируют входящий поток ионов калия через GIRK, активированный βγ-димером G-белка, и угнетают активность кальциевых каналов, что вызывает гиперполяризацию клетки. Именно таким образом устроены метаботропные ГАМК-рецепторы (гетеродимеры R1 и R2 субъединиц). Сходным образом устроен и 5-HT1A-рецептор.
Метаботропные ингибирующие рецепторы генерируют медленные тормозные постсинаптические потенциалы (длительностью от миллисекунд до минут). Они могут активироваться одновременно с ионотропными (с некоторыми типами ионотропных рецепторов они могут формировать «рецепторный дуплет» — гетеродимер) в одном и том же синапсе, что позволяет одному и тому же синапсу генерировать как быстрые, так и медленные тормозные потенциалы.
Формирование синапсов в ЦНС
Данные, полученные в исследованиях нервно-мышечных синапсов, можно попытаться экстраполировать на процессы синаптогенеза в ЦНС, даже несмотря на то, что исследования последних имеют намного более поздний характер. Эти два типа соединений имеют много общего. Оба типа синапса имеют консервативное базовое строение: пре-синаптическая нервная терминаль, синаптическая щель и принимающая постсинаптическая клетка. Оба типа синапса содержат высокую концентрацию везикул в активном центре, кластеры рецепторов на принимающей мембране, и, наконец, оба они изолированы специальными глиальными клетками. Процессы, происходящие в них, также достаточно консервативны. Это и кластеризация рецепторов, и локализованное повышение интенсивности синтеза белка в активных центрах, и синаптический прунинг путем элиминации малоактивных синапсов.
Несмотря на вышеобозначенные сходства, существует принципиальная разница между этими двумя соединениями
Поскольку постсинаптической клеткой уже не выступает миоцит, в корне отличаются действующие рецепторы и их нейротрансмиттеры. Что еще более важно, нейроны в пределах ЦНС иннервированы многими другими нейронами одновременно, и должны уметь интегрировать и обрабатывать большое количество информации на входе, когда как мышечные волокна иннервируются одним нейроном и сокращаются в ответ на одиночный импульс. В сочетании с пластичностью, характерной для нейронных связей, нейронная сеть в ЦНС имеет огромный потенциал для усложнения.
Сигналинг
Гомологом ацетилхолина и его рецепторов в ЦНС является глутамат и N-метил-D-аспартат-рецепторы (NMDA). Было показано, что активация NMDA-рецепторов инициирует каскад химических реакций, ведущих к формированию и стабилизации синапса. Повышенный уровень активности NMDA-рецепторов во время развития синапса позволяет увеличить приток кальция, который выступает в качестве вторичного сигнала. Помимо этого, факторами транскрипции активируются гены первичного ответа (ГПО), играющие важную роль в таких процессах, как, например, долговременная потенциация. Функция рецепторов NMDA также связана с эстрогеновыми рецепторами в гиппокампе. Эксперименты, проведенные с эстрадиолом, показывают, что воздействие эстрогена значительно увеличивает плотность синапсов и концентрацию белка.
Передача нервного импульса во время синаптогенеза зависит не только от активности отдельно взятого синапса, но и от окружающей этот синапс среды
Например, нейротрофический фактор головного мозга (BDNF) — продуцируется головным мозгом и регулирует несколько функций в развивающемся синапсе, включая усиление высвобождения нейротрансмиттеров, повышение концентрации везикул и биосинтез холестерина.
Холестерин является важной структурной составляющей липидного бислоя, выступающим как место действия самых разных сигнальных взаимодействий. Мутантные на наличие BDNF организмы обнаруживают значительные дефекты в росте нейронов и формировании синапсов. Помимо нейротрофинов, важную роль в процессе синаптогенеза играют также молекулы клеточной адгезии
Во многом синаптогенез зависит от корректного локального связывания одной адгезионной молекулы с другой. Показано, что дефекты в генах, кодирующих белки-нейролигины, одни из важнейших молекул клеточной адгезии, играющие роль в процессе стабилизации синапса, связаны с некоторыми случаями аутизма и умственной отсталости. Наконец, многие из этих сигнальных процессов могут регулироваться с помощью матриксных металлопротеиназ (ММП).
Морфология
Высокодинамичными сайтами, принимающими одновременно несколько входящих импульсов, являются дендритные шипики.
Этот морфологический динамизм обусловлен специфической регуляцией актинового цитоскелета, что, в свою очередь, позволяет регулировать образование синапсов. Морфологически, дендритные шипики можно разделить на три основных вида: филоподии, тонкие шипики и грибовидные шипики. Филоподии инициируют контакт с аксонами других нейронов. Филоподии новообразованных нейронов имеют тенденцию к образованию контактов с аксонами, иннервирующими множество синапсов, когда как филоподии более старых нейронов связываются преимущественно с аксонами, иннервирующими одиночный синапс. Благодаря высокой динамичности, одни типы шипиков способны переходить в другие, например, филоподии легко трансформируются в грибоподобные шипики, которые являются первичными сайтами глутаматных рецепторов.
Виды синапсов

Синапсы обеспечивают связи не только между нейронами головного мозга, но и с нервными клетками органов чувств, рецепторами, расположенными во внутренних органах, мышцах и связках. Поэтому существует большое разнообразие синапсов в зависимости от специализации нейронов, от характера их воздействия, от того белкового соединения, которое вырабатывается при прохождении импульса.
В нашей нервной системе существует два основных процесса, определяющих ее деятельность. Это возбуждение и торможение. В соответствии с ними и синапсы делятся на два вида:
- возбуждающие проводят сигналы, которые распространяют реакцию возбуждения нервных клеток;
- тормозящие обеспечивают прохождение нервного импульса, который передает нейронам «команду» торможения.
По месту расположения синапсы различаются:
- на центральные, расположенные в головном мозге;
- периферические, обеспечивающие связи нейронов за пределами мозга – в периферической нервной системе.
Передача импульсов через синаптическую щель тоже может проводиться разными способами, в соответствии с этим выделяют три вида синапсов:
- Химические синапсы расположены в коре головного мозга. Они проводят сигнал с помощью нейротрасмиттеров, которые образуются в результате биохимической реакции.
- Электрические – та часть синапсов, которые способны передавать электрический сигнал без посредников-медиаторов. Например, это касается нейронов, расположенных в зрительном рецепторе. В этом случае химическая реакция не происходит, и обмен сигналами осуществляется быстрее.
- Электрохимические синапсы сочетают в себе особенности обеих этих групп.
Также существует классификация синапсов по видам трансмиттеров. Например, если производится норадреналин, но синапсы эти называются адренергические, а если ацетилхолин, то – холинергические. Учитывая, что белков, вырабатываемых нейронами, несколько десятков видов, мы имеем очень объемную классификацию, которая здесь вряд ли уместна.
Мозг и потребности
Ключевая задача мозга — руководить поведением, которое в большинстве случаев нацелено на удовлетворение определенной нужды. Есть ряд базовых потребностей, с рождения встроенных в мозг и являющихся основой нашего поведения.
В перечень потребностей прежде всего входят витальные программы, ответственные за выживание человека: питание, защищенность, гомеостаз и так далее. Велика роль социальных программ, отвечающих за жизнь внутри сообщества. И есть особые программы, заставляющие стремиться к свободе, новизне, подражанию.
Центр каждой биологической потребности можно обнаружить в той или иной зоне мозга и проанализировать, на какие факторы реагирует этот центр. Как правило, значимы, во-первых, внешние сигналы, скажем, какие-то болевые стимулы. Во-вторых, внутренние сигналы, допустим, химический состав крови. Огромное значение для некоторых видов поведения имеет гормональный фон.
Врожденно заданная активность центра той или иной потребности во многом определяет наш темперамент, индивидуальную структуру нашей личности. Также очень значима генетическая основа и индивидуальный опыт, особенно тот, который приобретается в начале жизни, в ранний период онтогенеза. В зоопсихологии подобного рода события обозначают понятием «импринтинг».
Каждый конкретный поведенческий акт может приводить либо к удовлетворению потребности, либо к тому, что она не удовлетворяется. Если нужду удается удовлетворить, в мозге генерируются позитивные чувства. Они заставляют мозг запоминать успешные алгоритмы поведения. При фрустрации же возникают негативные чувства. На их основе происходит забывание, снижение рейтинга тех программ, которые окончились неудачей.
Кратковременная синаптическая пластичность
Значительное число различных форм кратковременной (от миллисекунд до нескольких минут) синаптической пластичности было обнаружено практически для всех синапсов организмов различного уровня сложности — от беспозвоночных до человека. Этот вид пластичности считается важным для кратковременной адаптации к сенсорной информации, изменений в поведении, а также кратковременной памяти.
Кратковременная пластичность инициируется короткими вспышками активности, которые вызывают временное накопление ионов кальция в пресинаптических терминалах. Вследствие этого накопления происходит изменение вероятности высвобождения нейротрансмиттера из-за непосредственной модуляции биохимических процессов, управляющих экзоцитозом.
Обработка парных импульсов — важнейшая задача кратковременной пластичности. Если клетка получает два импульса, разделённых коротким интервалом, ответ на второй импульс может быть как сильнее (потенциация), так и слабее (депрессия) ответа на первый. Ослабление парных импульсов наблюдается обычно при коротких интервалах времени между ними (меньше 20 мс); наиболее вероятной причиной такого ослабления может являться деактивация потенциал-зависимых натриевых и кальциевых каналов или временное уменьшение числа везикул в пресинаптическом терминале. При бо́льших интервалах между стимулами (20-500 мс) во многих случаях отклик на второй сигнал сильнее, чем на первый.
Проявление усиления или ослабления парных импульсов зависит от истории синапса. Эти формы пластичности сильно зависят от изменения вероятности высвобождения нейротрансмиттера в ответ на первый сигнал. Если эта вероятность высока, наблюдается тенденция к ослаблению второго сигнала; если же активация синапса одним сигналом маловероятна, разумно предположить, что следующий сигнал увеличит эту вероятность. Манипуляции, изменяющие вероятность отклика синапса на сигнал, могут изменить величину эффекта или даже изменить его характер — например, с усиления на ослабление.
Более долгоживущие формы пластичности возникают после серии стимуляций, произведённых с высокой частотой (судорожной, или тетанической стимуляции). Усиление (augmentation, facilitation) и пост-судорожная потенциация (post-tetanic potentiation, PTP) описывают увеличение выброса нейротрансмиттера продолжительностью от нескольких секунд (усиление) до нескольких минут (PTP). При этом также увеличивается вероятность выброса нейротрансмиттера за счёт накопления кальция в пресинаптическом терминале во время серии стимулов, которое может сочетаться с модификацией пресинаптических белков.
В некоторых случаях судорожная стимуляция приводит к депрессии синаптической связи, которая может продолжаться от нескольких секунд до минут. Обычно этот эффект возникает в синапсах с высокой вероятностью срабатывания, когда судорожная стимуляция приводит к быстрому опустошению запаса везикул или другим проявлениям ингибиторной машинерии.
Химия психики
Второстепенные медиаторы нужны для функционирования нашей психоэмоциональной сферы. К примеру, дофамин. С этой молекулой связана масса положительных эмоций. Нарушение функций дофамина приводит к таким патологиям, как паркинсонизм и шизофрения. Препараты, схожие с дофамином, работают как наркотики-психостимуляторы.
Еще один медиатор – серотонин. От него зависит целый ряд тормозных функций. Он контролирует центры негативных эмоций и уровень шума в мозговой коре. Благодаря серотонину мышление человека становится более чутким. С данным медиатором связаны препараты, которые мы относим к антидепрессантам. А еще на функции серотонина воздействуют наркотики, способные вызывать галлюцинации.
Эндорфины – ключевые медиаторы, связанные с контролем боли и опять-таки с центрами положительных эмоций. Поэтому на их основе созданы важнейшие группы анальгетиков, а также такие известные наркотические препараты, как морфин и героин, которые влияют на эндорфиновые синапсы.
Список медиаторов можно продолжить: аденозин, глицин, ацетилхолин, норадреналин… Любой из них крайне важен для функционирования мозга и внутренних органов. На их основе созданы важнейшие группы лекарств.
Строение нейронов
Схема нейрона
Тело клетки
Тело нервной клетки состоит из протоплазмы (цитоплазмы и ядра), ограниченной снаружи мембраной из липидного бислоя. Липиды состоят из гидрофильных головок и гидрофобных хвостов. Липиды располагаются гидрофобными хвостами друг к другу, образуя гидрофобный слой. Этот слой пропускает только жирорастворимые вещества (напр. кислород и углекислый газ). На мембране находятся белки: в форме глобул на поверхности, на которых можно наблюдать наросты полисахаридов (гликокаликс), благодаря которым клетка воспринимает внешнее раздражение, и интегральные белки, пронизывающие мембрану насквозь, в которых находятся ионные каналы.
Нейрон состоит из тела диаметром от 3 до 130 мкм. Тело содержит ядро (с большим количеством ядерных пор) и органеллы (в том числе сильно развитый шероховатый ЭПР с активными рибосомами, аппарат Гольджи), а также из отростков. Выделяют два вида отростков: дендриты и аксон. Нейрон имеет развитый цитоскелет, который проникает в его отростки. Цитоскелет поддерживает форму клетки, его нити служат «рельсами» для транспорта органелл и упакованных в мембранные пузырьки веществ (например, нейромедиаторов). Цитоскелет нейрона состоит из фибрилл разного диаметра: Микротрубочки (Д = 20—30 нм) — состоят из белка тубулина и тянутся от нейрона по аксону, вплоть до нервных окончаний. Нейрофиламенты (Д = 10 нм) — вместе с микротрубочками обеспечивают внутриклеточный транспорт веществ. Микрофиламенты (Д = 5 нм) — состоят из белков актина и, в отличие от других клеток, не содержат миозина, что делает невозможным сокращение в этих клетках, сами микрофиламенты особенно выражены в растущих нервных отростках и в нейроглии.(Нейроглия, или просто глия (от др.-греч. νεῦρον — волокно, нерв + γλία — клей), — совокупность вспомогательных клеток нервной ткани. Составляет около 40 % объёма ЦНС. Количество глиальных клеток в мозге примерно равно количеству нейронов).
В теле нейрона выявляется развитый синтетический аппарат, гранулярная эндоплазматическая сеть нейрона окрашивается базофильно и известна под названием «тигроид». Тигроид проникает в начальные отделы дендритов, но располагается на заметном расстоянии от начала аксона, что служит гистологическим признаком аксона. Нейроны различаются по форме, числу отростков и функциям. В зависимости от функции выделяют чувствительные, эффекторные (двигательные, секреторные) и вставочные. Чувствительные нейроны воспринимают раздражения, преобразуют их в нервные импульсы и передают в мозг. Эффекторные (от лат. effectus — действие) — вырабатывают и посылают команды к рабочим органам. Вставочные — осуществляют связь между чувствительными и двигательными нейронами, участвуют в обработке информации и выработке команд.
Различается антероградный (от тела) и ретроградный (к телу) аксонный транспорт.
Дендриты и аксон
Основные статьи: Дендрит и Аксон
Схема строения нейрона
Аксон — длинный отросток нейрона. Приспособлен для проведения возбуждения и информации от тела нейрона к нейрону или от нейрона к исполнительному органу.
Дендриты — короткие и сильно разветвлённые отростки нейрона, служащие главным местом для образования влияющих на нейрон возбуждающих и тормозных синапсов (разные нейроны имеют различное соотношение длины аксона и дендритов), и которые передают возбуждение к телу нейрона. Нейрон может иметь несколько дендритов и обычно только один аксон. Один нейрон может иметь связи со многими (до 20 тысяч) другими нейронами.
Дендриты делятся дихотомически, аксоны же дают коллатерали. В узлах ветвления обычно сосредоточены митохондрии.
Дендриты не имеют миелиновой оболочки, аксоны же могут её иметь. Местом генерации возбуждения у большинства нейронов является аксонный холмик — образование в месте отхождения аксона от тела. У всех нейронов эта зона называется триггерной.
Синапс
Основная статья: Синапс
Си́напс (греч. σύναψις, от συνάπτειν — обнимать, обхватывать, пожимать руку) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. Одни синапсы вызывают деполяризацию нейрона и являются возбуждающими, другие — гиперполяризацию и являются тормозными. Обычно для возбуждения нейрона необходимо раздражение от нескольких возбуждающих синапсов.
Термин был введён английским физиологом Чарльзом Шеррингтоном в 1897 г.
Мозг и мышление
Высшие функции больших полушарий связаны с ассоциативной корой. Ассоциативность здесь подразумевает то, что она объединяет многие информационные потоки. И на боковой поверхности полушарий мы видим прежде всего ассоциативную теменную кору и ассоциативную лобную кору. Первая занимает в основном заднюю часть теменной доли, располагается она между двумя главными сенсорными центрами. В итоге здесь собирается зрительная, слуховая, тактильная, вкусовая информация и прочие информационные потоки. Формируется целостная сенсорная картина внешнего мира.
Здесь возникает целостное восприятие некого объекта. Если это апельсин, то мы понимаем, что это фрукт, который называется «апельсин», что он имеет круглую форму, имеет оранжевый цвет, пахнет определенным образом и так далее. В этой зоне возникают подобного рода образы, и с ними у человека соединены словесные названия того или иного объекта окружающей среды. Именно ассоциативная теменная кора выступает местом локализации наших речевых центров. Когда этих центров становится много, они собираются в единую систему, которую можно назвать «информационной моделью внешнего мира». С помощью нее мы думаем, творим, мечтаем… Это очень важная часть нашей психической деятельности.
Лобная кора – это главный центр управления поведением. Здесь принимаются решения о запуске тех или иных программ. И первое, что она делает, — это оценивает выраженность различных потребностей. Этот участок мозга выбирает доминирующую нужду, а дальше он должен запустить программу, которая позволила бы эту нужду удовлетворить. При этом лобная кора учитывает сигналы от ассоциативной теменной коры, а также от центров памяти: от гиппокампа, от тех нейросетей, которые модифицировались в ходе долговременного обучения. Она запускает программу и мониторит ее реализацию. Такой мониторинг особенно важен в том случае, если программа длительная, если нужно за каждым этапом смотреть, удалось или не удалось достичь некой текущей цели.
Повреждение этого участка приводит к тому, что такие функции человеческого мозга, как воля и инициатива, очень сильно страдают. Кроме того, свойства ассоциативной лобной коры определяют такие особенности нашего темперамента, как импульсивность и настойчивость.
Примечания
- ↑ Purves et al. Neuroscience. 4th ed. Sunderland (MA): Sinauer Associates, Incorporated; 2008.
- Thompson S. M., Gahwiler BH. ACTIVITY-DEPENDENT DISINHIBITION .1. REPETITIVE STIMULATION REDUCES IPSP DRIVING FORCE AND CONDUCTANCE IN THE HIPPOCAMPUS INVITRO (англ.) // Journal of Neurophysiology (англ.)русск. : journal. — 1989. — Vol. 61. — P. 501—511.
- Levy et al. Principles of Physiology. 4th ed. (PA): Elsevier; 2005.
- Chavas J., Marty A. Coexistence of excitatory and inhibitory GABA synapses in the cerebellar interneuron network (англ.) // Journal of Neuroscience (англ.)русск. : journal. — 2003. — Vol. 23. — P. 2019—2031.
Передача возбуждения в синапсах
Механизм действия медиатора был изучен в серии работ английского ученого, лауреата Нобелевской премии Б. Катца и его сотрудников, которые в 1953 году обнаружили, что медиатор выделяется из нервных окончаний порциями. Каждая такая порция вызывает на мембране «отвечающей» клетки слабое изменение потенциала, которое обычно называют «миниатюрным потенциалом». Позднее с помощью электронного микроскопа, разделив предварительно синаптические нервные окончания на части, удалось выяснить, что медиатор в нервном окончании плотно упакован в маленькие синаптические пузырьки.
Множество таких пузырьков плавает внутри окончания. Когда пузырек прилипает к наружной мембране нервного окончания, содержимое такого пузырька — «квант» возбуждения — выделяется в синаптическую щель. Так возникает один миниатюрный потенциал. Кванты выделяются и в покое, но Катц показал, что нервный импульс в тысячи раз увеличивает среднюю их частоту, не меняя величину отдельной порции.
Но почему приходящий нервный импульс учащает слипание синаптических пузырьков с наружной мембраной нервного волокна? Сначала казалось, что причина очевидна. Нервный импульс, приходя к мембране, уменьшает на ней разность потенциалов. А еще раньше было известно, что в солевом растворе всевозможные воздействия, уменьшающие эту разность потенциалов, увеличивают частоту миниатюрных потенциалов. Но были поставлены другие опыты, в которых из наружного раствора удаляли ионы кальция, и тогда никакого увеличения частоты не наблюдалось, несмотря на приход нервного импульса. И наоборот, резко увеличить частоту выделения квантов можно безо всякого импульса — надо лишь поднять концентрации в наружном растворе любых непроникающих ионов или нейтральных молекул, например сахарозы.
Эта запутанная ситуация поставила в тупик английских исследователей. Распутать ее удалось авторам «кальциевой гипотезы» ученым Е. А. Либерману и его сотрудникам. Они исходили из простой физической идеи: для слипания пузырьков с наружной мембраной надо уменьшить поверхностный электрический заряд мембран — а это делают ионы кальция, которые входят в нервное окончание из-за того, что приходит нервный импульс. Поэтому-то в растворе, лишенном кальция, нет синаптической передачи.
Поскольку диаметр нервного окончания обычно меньше микрона, ввести туда ионы кальция для прямой проверки гипотезы нельзя. Но исследователи воспользовались тем, что кальций в нервных окончаниях всегда запасен в больших количествах внутри митохондрий, где его держит электрическое поле этих внутриклеточных «электростанций». Это позволило им, выбрасывая кальций из митохондрий различными агентами, снимающими электрическое поле, «ввести» кальций внутрь нервного окончания. Оказалось, что все вещества, снимающие электрическое поле на мембране митохондрий, резко повышают частоту выделения квантов ацетилхолина.
В дальнейшем эстафета вновь перешла к англичанам — им удалось поставить прямые опыты на гигантских синапсах кальмаров, в которых толщина нервного волокна достигает миллиметра. Так была подтверждена гипотеза Е. А. Либермана о роли кальция в синаптической передаче. (Однако каким образом пузырек раскрывается, чтобы излить свое содержимое в синаптическую щель, как его мембрана становится частью поверхностной мембраны нервного волокна — эти вопросы до сих пор не имеют ответа.).
После того как медиатор отработал, его нужно убрать. Для этого существуют разные механизмы. Один из них — ферменты-разрушители, молекулы, которые сидят в синаптической щели и разрезают медиатор на куски. Другой — клетка бережно собирает медиатор, чтобы использовать его вновь. Она втягивает внутрь себя часть мембраны, получившийся пузырек «отшнуровывается» и захватывает наружный раствор со всем его содержимым. Такой механизм удобен не только для клетки, но и для ученых, его изучающих. Используют его так: из обычного хрена выделяют фермент-разрушитель пироксидазу и вводят его в район синапса. Нервное окончание заглатывает пироксидазу вместе с медиатором. Нейроны, пославшие свои отростки в место, куда была введена пироксидаза, теперь легко обнаружить по специальной окраске, которая возникает из-за того, что пироксидаза, работая, расщепляет перекись водорода и выделяет молекулярный кислород. Так составляют карту связей между нейронами и разбирают принцип работы синапса.